Prophase
La prophase (du grec ancien πρό, pró : « avant ») est la première phase de la division cellulaire mitotique, comme méiotique chez les êtres vivants eucaryotes. Elle débute par la visualisation des chromosomes condensés (cette condensation de la chromatine en chromosomes se fait dès la phase G2, mais n'est pas discernable en microscopie optique) et se termine peu avant la dissociation de l’enveloppe nucléaire (caractéristique de la prométaphase). La prophase I est longue et complexe. Elle est formée de cinq stades : leptotène, zygotène, pachytène, diplotène et diacinèse.
Mitose

Cette phase débute par l'observation du matériel génétique (ADN), qui en temps normal est présent dans le noyau sous forme de chromatine décondensée, en structures très ordonnées, condensées et individualisées appelées chromosomes. Chaque chromosome est composé de deux chromatides reliés en partie par leur centromère. La prophase est caractérisée par la condensation des chromosomes et la formation du fuseau mitotique.
Les événements nucléaires
Pour les chromosomes, on assiste à la condensation progressive de leur chromatine. Les chromosomes sont formés de deux chromatides sœurs (possédant la même information génétique) qu’on ne peut pas encore visualiser totalement car reliées sur toute leur longueur par des complexes protéiques en forme d’anneau, les cohésines (qui assurent la cohésion des chromatides sœurs depuis la phase S jusqu’à la transition métaphase/anaphase). Ces cohésines interviennent lors de la méiose/mitose mais aussi pendant l'interphase, en jouant des rôles diversifiés (régulation de la transcription, réparation ADN). Jusqu'à la métaphase, les cohésines vont être remplacées par des condensines (responsables de la condensation) entraînant un condensation croissante et une association des chromatides sœurs décroissantes. Ces changements sont sous le contrôle du complexe CDK1-Cycline B1 (qui fixe les condensines) et de la kinase cdc5/Plk1 (qui enlève les cohésines).
Pour le nucléole, on voit un effacement progressif de tous les constituants nucléolaires (à l’exception du centre fibrillaire).
Quant à l'enveloppe nucléaire, elle prend un aspect ondulé dans certaines cellules. Après la prophase, donc pendant la prométaphase, la lamina nucléaire disparaît, entraînant la rupture de l’enveloppe nucléaire et sa dissociation en saccules (semblables à celles du réticulum endoplasmique).
Les événements cytoplasmiques
Un organite (la définition d'organite varie selon les auteurs) important de la prophase est le centrosome. Comme pour les chromosomes, le centrosome s'est dupliqué durant les phases S et G2 de l'interphase, avant le début de la prophase. Au début de la prophase le centrosome contient quatre centrioles, mais va se diviser en deux pour donner deux centrosomes à deux centrioles. Chacun des deux centrosomes migre vers un pôle de la cellule en glissant le long de l’enveloppe nucléaire. Dès la phase suivante, ils seront en position diamétralement opposée, formant les deux pôles de la cellule. Certaines cellules eucaryotes, notamment les cellules végétales, sont dépourvues de centriole.
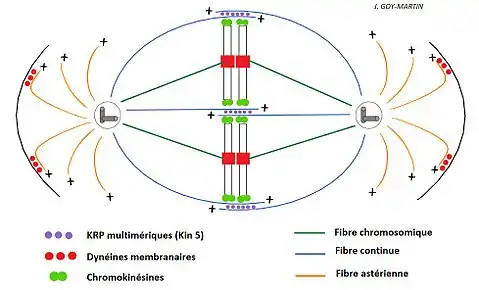
Le cytosquelette de microtubules se réorganise pour former le fuseau mitotique, structure bipolaire qui s'étend entre les deux centrosomes et reste à l'extérieur du noyau (dans le cytosol). Ce fuseau mitotique prophasique se décompose en fibres astériennes et fibres continues.
- Les fibres astériennes se retrouvent autour de chaque centrosome. Ce sont des fibres formées de faisceaux de microtubules qui partent en rayons, avec un aspect étoilé.
- Les fibres continues (ou microtubules chevauchants) permettent la croissance du fuseau mitotique, et donc la migration des centrosomes. On observe une extrême instabilité de ces microtubules (le complexe CDK1-Cycline B1 phosphoryle différentes MAP (MT Assocated Protein), compensée par une augmentation de la capacité de nucléation des centrosomes (augmentation du nombre de ƔTuRC). Les Kin 5 (ou Kinésine 5 ou Kinésines related protein) multimériques sont responsables de la séparation des centrosomes en marchant vers l’extrémité + des microtubules. Notons que ces fibres sont qualifiées de "continues" à cause d'une illusion d'optique présente lors de leur observation par microscopie optique (mise à jour par la microscopie électronique).
Méiose
Stades
La première division de la méiose se divise en 5 stades différents :
- Leptotène
- (du grec : leptos = mince et tainia = ruban)
On voit apparaître les chromosomes individualisés dont les télomères commencent à s'attacher à l'intérieur de la membrane nucléaire. Les chromatides sœurs appariées commencent à se condenser sous forme de boucles bordant un élément protéique axial dense.
- Zygotène
- (du grec zygon = joug et tainia = ruban) :
Les homologues se rapprochent selon un processus appelé synapsis, qui est intimement lié à l'assemblage du complexe synaptonémal. Au début du stade zygotène, les télomères des chromosomes sont regroupés au niveau d'un point sur l'enveloppe nucléaire ce qui fait apparaître l'arrangement en bouquet des chromosomes.
- Pachytène
- (du grec pakhus = épais) :
La synapsis est terminée, les homologues sont reliés entre eux par l'intermédiaire du complexe synaptonémal.
- Diplotène
- (du grec diploos = double) :
Au début de ce stade, le complexe synaptonémal se désintègre et les chromosomes se décondensent. Plus tard, ils se condensent à nouveau, les chromatides-sœurs restent étroitement associées tandis que les chromosomes homologues tendent à se séparer les uns des autres et ne sont plus retenus ensemble que par l'intermédiaire des chiasmas. Ce stade peut durer des jours ou même des années, selon le genre d'organisme et de son type (ovocytes de 1er ordre dans l'ovaire bloqués dans le stade diplotène jusqu'à leur transformation en ovocytes de 2e ordre, c'est-à-dire de la vie fœtale à chaque cycle adule chez la femelle). La transcription des chromosomes est très active au cours du stade diplotène, notamment chez les femelles car l'œuf se consacre activement au stockage du matériel qui sera utilisé dans les toutes premières divisions du développement embryonnaire.
- Diacinèse
- (dia = à travers et kinesis = mouvement) :
Au cours de ce stade, les chromosomes homologues raccourcissent et se condensent une fois de plus. La plaque de rattachement qui retenait les chromosomes homologues à la paroi du noyau disparaît tandis que les chromosomes s’épaississent.