Fuseau mitotique
Le fuseau mitotique, ou appareil mitotique achromatique, est un système mis en place par les cellules eucaryotes pour permettre la migration des chromatides lors de la division cellulaire dès le stade de la prophase. Il est constitué de microtubules et de protéines associées et forme un véritable fuseau entre les pôles opposés d’une cellule. Il assure une bonne répartition des chromosomes dans les cellules lors de la division. Il est donc indispensable à un bon développement cellulaire.
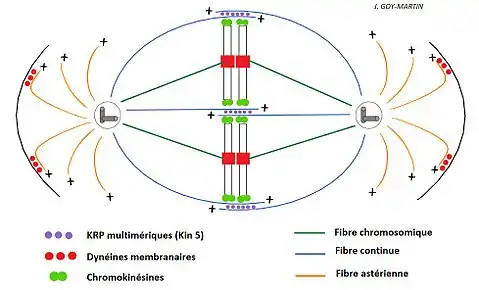
Notons que certains auteurs distinguent le fuseau mitotique (seulement les fibres chromosomiques et continues) et l'appareil achromatique (fuseau + centrosomes + fibres astériennes).
Les différents types de fibres
Les fuseaux mitotiques sont constitués de microtubules qui se polymérisent à partir du centrosome. Dans la cellule en division, il existe trois classes différentes de fibres. Bien qu'elles soient de constitution similaire, leurs rôles divergent.
Ces fibres se développent autour du centrosome ou microtubule organizing center (MTOC) et se dirigent vers la membrane cytoplasmique de la cellule. L'hypothèse émise est qu'elles jouent un rôle dans la mise en place du fuseau et interviendraient dans un mécanisme biologique survenant à la suite de la mitose, c'est la cytodiérèse. Après la cytodiérèse, les fibres d'asters serviraient à la formation du cytosquelette de chaque cellule fille.
- Fibres polaires ou continues ou chevauchantes :
Ces fibres vont se développer à partir du centrosome ou microtubule organizing center (MTOC) pour rejoindre le centre de la cellule en mitose et s'insérer (s'intercaler) entre les autres fibres polaires provenant du deuxième centrosome localisé à l'autre pôle de la cellule. Elles se lient grâce à des protéines motrices dont la chromokinésine qui se situe sur les télomères des chromosomes. En aucun cas elles ne se prolongent sur une longueur suffisante pour atteindre le centrosome du pôle opposé.
- Fibres chromosomiques ou kinétochoriennes :
Ces fibres se polymérisent du centrosome vers les chromosomes. Lors de la prométaphase, l'extrémité + du fuseau kinétochorien se fixe sur un complexe protéique situé au niveau du centromère des chromosomes. La fixation de ce complexe est faite par des protéines motrices, dont la dynéine et la kinésine.
Dynamisme moléculaire
Assemblage et désassemblage du fuseau
Le fait que les fuseaux mitotiques soient constitués de microtubules en fait une structure cellulaire dynamique qui se polymérise et se dépolymérise à tout moment. La longueur des fuseaux varie selon la polymérisation ou la dépolymérisation des microtubules pendant la mitose. Ces deux mécanismes sont dus à un flux des sous-unités d'alpha et beta tubuline. Il y a selon les besoins cellulaires des ajouts très rapides de sous-unités de tubuline alpha ou bêta permettant aux fuseaux de se développer suffisamment pour se fixer soit sur les kinétochores, soit sur des fibres polaires opposées. Lorsqu'il y a dépolymérisation du microtubule, cela se traduit par une perte de sous-unités de tubuline, ce qui entraîne un raccourcissement des fibres. Même si l'ajout et la soustraction de sous-unités de tubuline se font aux deux extrémités des fibres, les extrémités - restent toujours vers le centrosome, ce qui permet de déplacer le chromosome ou la chromatide attaché aux extrémités + des fibres. La vitesse de polymérisation/dépolymérisation des fuseaux est fonction de son extrémité : à l'extrémité + elle sera rapide tandis qu'à l'extrémité - elle sera lente.
Mitose
Au début de la mitose, les deux centrosomes se séparent, et chacun forme le noyau d'un arrangement de microtubules en rayons appelés aster. Les deux centrosomes se déplacent vers les deux côtés opposés du noyau pour former les deux pôles du fuseau mitotique. Lors de la prophase, les chromosomes répliqués se condensent et les fuseaux mitotiques commencent à s'assembler à l'extérieur du noyau. Pendant la pro-métaphase, l'enveloppe nucléaire se rompt, ce qui permet aux fuseaux d'entrer en contact avec les chromosomes et de les lier par l'intermédiaire de leurs kinétochores. L'attachement des microtubules sur les kinétochores se fait à la partie positive des microtubules. Durant la métaphase, le fuseau mitotique rassemble tous les chromosomes au niveau de son centre (plaque équatoriale). Pendant l'anaphase, les deux chromatides sœurs de chaque chromosome répliqué se dissocient de façon synchrone, et le fuseau chromosomique les tire en directions opposées vers les deux pôles de la cellule. Au cours de la télophase, les fibres polaires sont à l'origine d'une élongation de la cellule permettant de faire de la place pour les deux cellules filles à venir. Le fuseau mitotique permet la séparation des chromosomes et participe à la formation des deux cellules filles.
Régulation du fuseau
Il existe dans la cellule un équilibre entre la tubuline sous forme d'hétérodimères et la forme polymérisée des microtubules. Cet équilibre est extrêmement sensible, par exemple aux variations de la concentration de calcium. Comme la cellule est capable, grâce à des systèmes tampons et des pompes, de moduler localement et très rapidement la concentration de calcium, elle dispose donc d'un mécanisme de régulation très efficace et elle a la possibilité de contrôler la formation de microtubules et leur dissociation en dimères ou en monomères de tubulines. Un deuxième mécanisme de contrôle, qui est cependant à plus long terme, est la stabilisation des microtubules par des protéines associées ou des modifications chimiques comme une acétylation ou une détyrosylation.
Les fuseaux mitotiques subissent deux types de contrôles.
- L'origine du premier est une protéine située dans le kinétochore de chaque chromatide. Il a été mis en évidence que la protéine Mad 2 (mitotic arrest deficient 2) retarde la progression de la mitose lorsque certains chromosomes ne sont pas alignés dans la plaque métaphasique. Mad 2 empêche des réactions protéolytiques qui permettent à la cellule de rentrer dans le stade de l'anaphase. Quand les chromosomes sont alignés dans la plaque métaphasique, les kinétochores perdent cette protéine. La disparition de Mad 2 permet la suite de la mitose.
- Le deuxième est un contrôle mécanique, c'est le type de tension que subit le chromosome qui lui permet d'émettre ou non un signal d'inhibition qui retarde l'anaphase. La tension mécanique présente lorsque le chromosome est attaché uniquement à un microtubule chromosomique permet l'émission du signal d'inhibition. Lorsque chaque chromatide du chromosome est reliée à un microtubule, alors la tension mécanique change et le signal d'inhibition est stoppé. Ce sont ces deux systèmes qui régulent l'entrée en anaphase de la cellule.
Poisons des fuseaux mitotiques
Il existe un certain nombre de molécules chimiques qui peuvent être considérées comme des poisons du fuseau mitotique. C’est le cas des taxanes, de la colchicine et des vinca-alcaloïdes. Ces molécules bloquent la polymérisation ou la dépolarisation des microtubules. Certaines de ces molécules sont utilisées sur le plan thérapeutique pour soigner les tumeurs cancéreuses à fort index prolifératif. Il est donc possible de ralentir leur progression ou de les détruire en bloquant le fuseau mitotique et donc la mitose des cellules tumorales en évitant de toucher les cellules normales qui prolifèrent beaucoup moins.
Notes et références
- B. Alberts, A. Jonhson, J. Lewis, M. Raff, Biologie moléculaire de la cellule 4e édition, Flammarion Médecine-Sciences, Traite de médecine, 09/07/2004, (ISBN 2-257-16219-6), p. 1031
- G. Karp, Biologie cellulaire et moléculaire 2e édition, De Boeck, 01/04/2004, (ISBN 2-8041-4537-9), p. 593:604
- H.Plattner, J. Hentschel, Manuel de poche de Biologie cellulaire 3e édition, Flammarion Médecine-Sciences, atlas de poche, 01/04/2009, (ISBN 978-2-257-00004-0), p. 284, 285, 397
- B. Alberts, D. Bray, K. Hopkin, A. Jonhson, J. Lewis, M. Raff, K. Roberts, P. Walter, L'essentiel de la biologie cellulaire 2e édition, Flammarion Médecine-Sciences, Traite de médecine, 19/08/2005, (ISBN 2-257-15123-2), p. 640:655