Apatosaurus
Apatosaurus est un genre éteint de dinosaures herbivores sauropodes géants de la famille des diplodocidés. Il a vécu en Amérique du Nord au Jurassique supérieur (Kimméridgien et Tithonien), il y a environ entre 156,3 et 146,8 Ma (millions d'années)[3] - [4]. Leurs restes fossiles ont été découverts dans les États de l'ouest et du centre des États-Unis dans la formation de Morrison.

Espèces de rang inférieur
- † Apatosaurus ajax Marsh[1], 1877, espèce type
- † Apatosaurus louisae Holland[2], 1916
Étymologie
Le nom de genre Apatosaurus vient des mots du grec ancien apatē ( ἀπάτη) / apatēlos ( ἀπατηλός ) signifiant « déception » / « trompeur », et « sauros » ( σαῦρος) signifiant « lézard » pour donner « lézard trompeur ». Marsh lui a donné ce nom car les arcs hémaux de l'animal ressemblaient plus à ceux d'un mosasaure, un grand reptile marin, qu'à ceux d'un dinosaure[1].
Apatosaurus et Brontosaurus
L'apatosaure est encore souvent désigné, par erreur, dans le grand public, comme brontosaure ou Brontosaurus (du grec signifiant littéralement « lézard tonnerre »), genre qui a été considéré dès 1903 comme un synonyme junior d'Apatosaurus.
Cependant en 2015, une synthèse sur les Diplodocidae réalisée par Emanuel Tschopp, Octavio Mateus et Roger B.J. Benson en 2015 propose de réhabiliter le genre Brontosaurus auquel ils affilient trois espèces[5], ce qui n'est pas accepté par de nombreux paléontologues[6].
Création des deux genres
La confusion vient du fait que le paléontologue américain Othniel Charles Marsh qui a découvert les premiers fossiles leur a attribué le nom Apatosaurus en 1877[1] mais, deux ans après cette découverte, Marsh découvrit les restes d'un autre individu, un juvénile, qu'il n'identifie pas comme appartenant au même genre et qu'il nomme Brontosaurus[7].
Mise en synonymie sous le nom d'Apatosaurus
En 1903, un autre paléontologue américain, Elmer Samuel Riggs, réétudia les restes des deux spécimens et conclut qu'en réalité ils devaient être rangés dans le même genre[8]. Le genre Apatosaurus ayant été nommé en premier, le principe d'antériorité du code international de nomenclature zoologique (CINZ) lui donne la priorité, en reléguant Brontosaurus au rang de simple synonyme[8].
Résilience du terme brontosaure
Le terme de brontosaure est resté cependant courant dans la littérature non-scientifique, ainsi que dans les films, et bien connu du grand public, pour qui il apparaît même parfois comme un synonyme du mot dinosaure. Le service postal gouvernemental des États-Unis (United States Postal Service) imprime même en 1989 des timbres figurant des dinosaures incluant Brontosaurus aux côtés de Tyrannosaurus, Stegosaurus, Pteranodon, ce dernier n’étant d'ailleurs pas un dinosaure. Cette usage tardif du terme Brontosaurus à la place de celui d'Apatosaurus soulève une polémique de scientifiques qui critiquent l'institution comme « favorisant l'analphabétisme scientifique »[9]. Le célèbre paléontologue américain Stephen Jay Gould en fera même le sujet d'un de ses livres en 1991 « Bully for Brontosaurus » (« La Foire aux dinosaures : réflexions sur l’histoire naturelle »), où il reconnait l'importance du mot Brontosaurus dans l'imagination populaire, même si, d'un point de vue scientifique il ne peut qu'être d'accord avec la mise en synonymie de Riggs en 1903[10].
Tentative de réhabilitation scientifique du genre Brontosaurus
En 2015, Emanuel Tschopp, Octavio Mateus et Roger B.J. Benson dans leur étude des Diplodocidae concluent qu'Apatosaurus excelsus était en fait suffisamment différent de l'espèce type du genre Apatosaurus, pour conserver son attribution originale de Brontosaurus excelsus (Marsh, 1879)[5]. Leur étude s'appuie sur l'analyse de 81 spécimens sur lesquels ils comparent les variations de 477 caractères morphologiques. Brontosaurus se distinguerait d'Apatosaurus par[5] :
- un cou moins large, supposé moins fort ;
- des vertèbres dorsales postérieures présentent un corps vertébral plus long que large ;
- une omoplate dont la partie arrière jusqu'au bord acromial et à la lame distale présente un sillon ;
- le bord acromial de la lame scapulaire distale porte un renflement arrondi ;
- un rapport de la longueur/largeur de l'os astragale supérieur à 0,55.
Cette proposition qui va à l'encontre des règles habituelles de la taxonomie a ouvert une polémique[11].
Des experts comme Paul Barrett, responsables des dinosaures au Musée d'histoire naturelle de Londres, semblent dépités : « Ça nous a pris beaucoup de temps pour convaincre les gens qu'ils ne doivent pas utiliser le nom brontosaure. Et maintenant que l'on y est arrivé, il semble qu'il va falloir revenir en arrière et dire "En fait, vous pouvez le faire" »[6].
Description
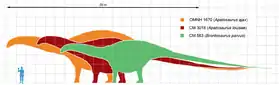
Apatosaurus ajax (orange), Apatosaurus louisae (rouge),
avec un humain (bleu) et Brontosaurus parvus (vert).

sculpté en 1931,
pour être monté sur un squelette post-crânien d'Apatosaurus.
Muséum d'histoire naturelle Peabody.
Taille
L'apatosaure est célèbre pour sa longue colonne vertébrale, son long cou, et sa queue effilée qui comptait 82 os ; c'est un très grand sauropode et donc l'un des plus grands animaux terrestres ayant jamais vécu.
La plupart des estimations de taille sont basées sur un spécimen, l'espèce type d'Apatasaurus louisae du Carnegie Museum of Natural History de Pittsburgh, référencé CM 3018. En 1936, sa colonne vertébrale a été mesurée à 21,8 mètres par Charles Gilmore[12]. En 2015, sa masse est estimée entre 21,8 et 38,2 tonnes, similaire à celle du genre Dreadnoughtus[13].
Quelques spécimens de l'espèce A. ajax, comme celui du Muséum d'histoire naturelle d'Oklahoma, référencé OMNH 1670, correspondent à des individus qui seraient 11 à 30 % plus longs que CM 3018, soit 24 à 28 mètres (Thomas Holtz retient la longueur de 26 mètres en 2011[14]), suggérant une masse près de deux fois plus importantes que celle de CM 3018, soit 32,7 à 72,6 tonnes[15].
Crâne

spécimen CMC VP 7180.
Le crâne d'Apatosaurus n'a pas été découvert lors des premières descriptions de l'animal. Ainsi plusieurs squelettes post-crâniens d'Apatosaurus ont été remontés avec des crânes appartenant aux genres Camarasaurus ou Brachiosaurus.
Le premier crâne, celui d'Apatosaurus louisae, ne fut découvert qu'en 1909 à quelques mètres des os du cou ; il est très similaire à celui de Diplodocus[16]. Cependant, malgré cette découverte, le paléontologue Henry Fairfield Osborn et certains de ses collègues refusent cette association et c'est ainsi que la plupart des squelettes d'Apatosaurus continuent à porter des crânes ne leur appartenant pas ou avec des crânes hypothétiques en plâtre jusque dans les années 1970. En 1975, John Stanton McIntosh et David Berman redécrivent les crânes connus de Diplodocus et d'Apatosaurus et confirment qu'Apatosaurus (et Brontosaurus) possédaient un crâne ressemblant à celui des Diplodocus. Ils réattribuent de nombreux crânes supposés appartenir au genre Diplodocus au genre Apatosaurus[17].
Comme chez la plupart des sauropodes le crâne est relativement petit par rapport à la taille de l'animal. Les mâchoires sont garnies de dents spatulées (en forme de ciseau) adaptées à un régime herbivore[18]. Le museau d'Apatosaurus, comme celui des diplodocoïdes proches est de forme carrée, sans toutefois être aussi géométrique que le rebbachisauridé Nigersaurus[19].
Colonne vertébrale

Muséum d'histoire naturelle Peabody, (YPM 1860)
en vue latérale et antérieure.
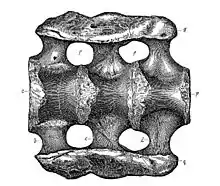
La formule vertébrale de l'holotype d'A. louisae est de 15 cervicales, 10 dorsales, 5 sacrées et 82 caudales. Le nombre de ces dernières varie cependant même à l'intérieur d'une même espèce[12].
Comme chez les autres sauropodes les vertèbres cervicales sont profondément divisées (bifurquées) ; elles portent des épines neurales séparées par un large sillon qui donne un cou très large à l'animal[18].
Les vertèbres cervicales d'Apatosaurus et de Brontosaurus sont plus larges et plus robustes que celles des autres diplodocidés ; elles ressemblent à celles de Camarasaurus[12] - [20]. De plus, elles portent des côtes cervicales plus allongées vers le sol que chez les diplodocinés. Le cou est cependant allégé par une grande pneumatisation des vertèbres cervicales qui renferment de nombreux sacs à air. Chez Apatosaurus louisae, le couple atlas-axis des premières cervicales est presque fusionné. Les 10 côtes dorsales sont par contre articulées librement aux vertèbres[12].
La forme de la queue est inhabituelle pour un diplodocidé, car la hauteur des processus épineux des vertèbres décroît rapidement depuis les hanches pour donner une queue relativement fine. Comme chez les autres diplodocidés, la queue de ce quadrupède ressemble dans sa partie finale à un fouet[12].
Membres
Les membres d'Apatosaurus sont très robustes avec les pattes avant un peu plus courtes que les postérieures[21].
La ceinture pelvienne montre des ilions robustes et des pubis et ischions fusionnés.
Les fémurs d'Apatosaurus sont très solides et sont parmi les plus robustes chez les sauropodes. Les os du tibia et du péroné sont différents des os élancés de Diplodocus, mais sont presque indiscernables de ceux de Camarasaurus. Le péroné est plus long et plus mince que le tibia[12] - [22].
Chaque patte avant portait une seule grande griffe, un trait partagé par tous les sauropodes plus évolués que le genre Shunosaurus[12] - [22]. Cette phalange unguale, en forme de griffe, est légèrement courbée et tronquée à son extrémité. Par contre, les trois premiers orteils (vers l'intérieur) possèdent des griffes sur chaque membre postérieur[23].
Paléobiologie
Au cours du XIXe siècle et au début du XXe siècle, les sauropodes étaient supposés être trop massifs pour soutenir leur propre masse à terre[24]. Il avait été théorisé qu'ils ne pouvaient vivre qu'en partie immergés dans l'eau. Ces théories ont été réfutées au cours du XXe siècle et au début du XXIe siècle, et les sauropodes sont aujourd'hui considérés comme des animaux franchement terrestres[25].
La morphologie du museau et des dents, disposées de façon rectiligne, plaide pour un régime herbivore non sélectif de plantes à hauteur du sol[19]. Des analyses de la morphologie des vertèbres du cou de l'animal ont montré que, contrairement à l'image classique d'animaux au cou redressé pour attraper de la végétation à grande hauteur, celles-ci n'étaient pas très flexibles et qu'Apatosaurus était adapté pour consommer une végétation basse[26] - [27] - [28]. Cette hypothèse est toutefois remise en cause en 2014 par Taylor qui, après avoir étudié la quantité et la position des cartilages intervertébraux, conclut à l'opposé que le cou d''Apatausaurus, comme celui de Diplodocus était très flexible[29]. Stevens et Parrish en 1999 estiment qu'Apatosaurus pouvait manger des fougères, des Bennettitales, ptéridospermes ou « fougères à graines », des Equisetales (sortes de prêles) et des algues[26]. Ces mêmes auteurs en 2005 envisagent qu'il se nourrissait de plantes sub-aquatiques à partir des rives[28].
Une étude en 2015 de Taylor et ses collègues sur les cous d'Apatosaurus et Brontosaurus montrent qu'ils sont très différents de ceux des autres diplodocidés et que cette différence serait due à l'utilisation, par ces deux genres, de leur cou comme d'une arme dans des combats lors de compétitions intraspécifiques[20].
L'étude des pistes fossiles des sauropodes comme Apatosaurus et Brontosaurus, démontrerait à partir de l'analyse des leurs empreintes de pas, qu'ils pouvaient marcher entre environ 25 et 40 kilomètres par jour et que leur vitesse maximale pouvait atteindre de 20 à 30 kilomètres par heure[18].
La morphologie de la queue d’Apatosaurus a donné lieu à des interrogations sur sa capacité à émettre des claquements comparables à ceux d'un fouet, claquements qui sont émis par l'extrémité de l'arme lorsqu'elle franchit le mur du son. L'hypothèse est avancée pour la première fois en 1997 par Nathan Myhrvold, un employé de Microsoft passionné de paléontologie, sur la base d'une simulation par ordinateur qu'il avait co-élaborée avec le paléontologue canadien Philip John Currie[30] - [31]. L'hypothèse est accueillie avec curiosité par plusieurs paléontologues. D'autres sont plus sceptiques, comme Kenneth Carpenter[32]. Après de nouvelles recherches, Myrhold présente une maquette de queue de sauropode lors de la conférence de la Society of Vertebrate Paleontology en 2015 à l'appui de son hypothèse. En 2022, de nouvelles recherches menées par Simone Conti de l'École des sciences et des technologies de l'Université nouvelle de Lisbonne, au Portugal, élaborent des simulations informatiques plus précises en prenant en compte un plus grand nombre de facteurs. Elles concluent que les sauropodes avaient des queues bien moins souples qu'on ne le pensait auparavant et que l'extrémité de la queue ne se déplaçait pas à plus de 33 mètres par seconde au maximum, soit bien moins vite que ne le supposaient les simulations de Myrhold (et bien en deçà de la vitesse nécessaire pour franchir le mur du son). De plus, toujours selon ces simulations, les tissus organiques du bout de la queue n'auraient pas supporté d'atteindre des vitesses pareilles[31].
Paléoécologie

Muséum d'histoire naturelle d'Oklahoma.

Les fossiles d'Apatosaurus proviennent de la formation géologique de Morrison. Ce sont des sédiments déposés dans un environnement alluvial à marin peu profond. Leur datation radiométrique indique un âge entre 156,3 et 146,8 Ma (millions d'années), couvrant à peu près tout l'intervalle du Kimméridgien et du Tithonien, les deux derniers étages du Jurassique supérieur[3] - [4]. Cette formation, caractéristique du bassin de Morrison, qui s'étend du Nouveau-Mexique jusqu'à l'Alberta et la Saskatchewan, s'est déposée dans un environnement semi-aride alternant des saisons sèches et humides. Les sédiments en provenance de bassins de drainages par des rivières se sont déposés dans des zones basses marécageuses, de lacs, de chenaux et de plaines alluviales[33] - [34] - [35].
Le registre fossile de la formation de Morrison est dominé par les sauropodes géants[36]. Il s'agit de Camarasaurus, Apatosaurus, Brontosaurus, Diplodocus et Brachiosaurus. Ils sont associés à des ornithischiens : Camptosaurus, Dryosaurus et Stegosaurus. Les théropodes carnivores associés sont Allosaurus, Ceratosaurus, Ornitholestes, Saurophaganax et Torvosaurus[37].
Les autres vertébrés qui partagent ce paléo-environnement sont des poissons à nageoires rayonnées, des grenouilles, des salamandre (amphibien)s, des tortues, des sphénodontes, des lézards, terrestres et aquatiques, des crocodylomorphes, et plusieurs espèces de ptérosaures. Des coquilles de bivalves et d'escargots aquatiques sont aussi courantes. La paléoflore se compose d'algues vertes, de champignons, de mousses, de « prêles », cycadophytes, de ginkgos et de plusieurs familles de conifères[38].
Apatosaurus est le second sauropode le plus abondant dans cet écosystème de la formation de Morrison après Camarasaurus[36] ; ses fossiles sont cependant plus concentrés dans la partie supérieure (tithonienne) de la formation et il s'agit presque exclusivement de restes d'A. ajax, car ceux d'A. louisae sont rares.
Malgré sa taille importante, Apatosaurus n'est pas le plus grand sauropode découvert dans cette formation, il était surpassé par Supersaurus[21].
Inventaire des fossiles retrouvés
Des fossiles de ce dinosaure ont été trouvés en relativement grande quantité (une douzaine de squelettes) en Amérique du Nord : aux États-Unis à Nine Mile Quarry et Bone Cabin Quarry au Wyoming, et sur des sites du Colorado, de l'Oklahoma, de l'Utah ; ce sont, avec leurs attributions originales :
- A. ajax : YPM 1860, 1861, 1840, 4676 - AMNH 460 - NSMT-PV 20375
- A. louisae : CMNH 3018, 3378, 11162 - LACM 52844
- « A. excelsus » : YPM 1980, 1981
- « A. minimus » : AMNH 675
Les différentes espèces
Espèces valides
Seules deux espèces d'apatosaures sont considérées comme valides, A. ajax et A. louisae.
Historique
De nombreuses espèces d'Apatosaurus ont été créées à partir de fossiles très partiels. Marsh dans le contexte très singulier de la guerre des os avec Edward Drinker Cope a nommé autant d'espèces qu'il le pouvait, la plupart basées sur des fossiles fragmentaires non diagnostiques.
En 2005, Paul Upchurch et ses collègues publient une synthèse sur le genre Apatosaurus. Ils en concluent qu'A. louisae est l'espèce la plus basale, suivie par le spécimen FMNH P25112, puis une polytomie constituée d'A. ajax, A. parvus et A. excelsus[23].
En 2015, leur analyse a été révisée et élargie par l'intégration de nombreux autres spécimens de diplodocidés, par Tschopp, Mateus et Benson qui ne retiennent que deux espèces d'Apatosaurus (A. ajax et A. louisae), et différencient le genre Brontosaurus, qu'ils réhabilitent, du genre Apatosaurus[5].
L'analyse des différents spécimens de diplodocidés étudiés et leur attribution aux genres et espèces considérés comme valides dans cette étude est résumée dans le cladogramme ci-dessous[5] :
| Apatosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Apatosaurus ajax
L'espèce a été nommée par Marsh en 1877 en référence à Ajax, le héros de la mythologie grecque[1] - [39]. Marsh a choisi comme holotype un squelette incomplet d'un spécimen juvénile (YPM 1860). Cette espèce a été moins étudiée que Brontosaurus et A. louisae, car les fossiles de cet holotype étaient trop incomplets.
En 2005, Paul Upchurch assigne plusieurs autres spécimens à l'espèce : YPM 1840, NSMT-PV 20375, YPM 1861 et AMNH 460[23].
En 2015, Emanuel Tschopp et ses collègues ne retiennent que l'holotype YPM 1860 comme appartenant sans conteste à l'espèce A. ajax. Le spécimen AMNH 460 appartiendrait, soit au genre Brontosaurus, soit à un éventuel nouveau taxon. Par ailleurs, YPM 1861 et NSMT-PV 20375 ne montrent que peu de caractères différents et ne peuvent être distingués d'A. ajax. Comme YPM 1861 est l'holotype d'« Atlantosaurus » immanis, il deviendrait un synonyme junior d'A. ajax[5].
Apatosaurus louisae

L'espèce a été nommée par William Jacob Holland en 1916, à partir d'un squelette partiel découvert en Utah[40]. L'holotype est référencé CM 3018. D'autres spécimens sont attribués à l'espèce : CM 3378, CM 11162 et LACM 52844. Les deux premiers sont représentés par des colonnes vertébrales, tandis que le dernier est constitué d'un crâne et d'un squelette presque complet.
En 2015, E. Tschopp et ses collègues concluent que le spécimen type d'Apatosaurus laticollis (YPM 1861) est très proche de CM 3018, et qu'il devient ainsi un synonyme junior d'A. louisae[5].
Espèces réattribuées

soit à A. ajax, soit à une nouvelle espèce plus basale.

au Muséum américain d'histoire naturelle,
tel que remonté en 1995.

Apatosaurus sp. (BYU 17096),
surnommé « Einstein ».
- Apatosaurus grandis est nommé en 1877 par Marsh dans l'article décrivant A. ajax. La description et la figuration sont sommaires[12]. Un an plus tard, Marsh dira que l'attribution au genre Apatosaurus était provisoire et le spécimen est réattribué à un nouveau genre Morosaurus[41]. Depuis Morosaurus a été mis en synonymie du genre Camarasaurus et plus précisément de C. grandis[36].
- Apatosaurus excelsus est à l'origine l'espèce type du genre Brontosaurus (B. excelsus), créé par Marsh en 1879[7]. En 1903, Elmer Riggs reclasse Brontosaurus comme un synonyme junior d'Apatosaurus, l'espèce devenant A. excelsus[8]. En 2015, Tschopp, Mateus et Benson font valoir que lors de sa première description par Marsh, l'espèce était suffisamment distincte d'Apatosaurus pour mériter d'être érigée comme un nouveau genre ; ils lui redonnent donc son nom original : Brontosaurus excelsus[5].
- Apatosaurus parvus, d'abord décrit en 1902 par Peterson et Gilmore comme un spécimen juvénile du genre Elosaurus[42], a été attribué à Apatosaurus in 1994, et puis en 2015 à Brontosaurus en compagnie de plusieurs autres spécimens adultes[5].
- Apatosaurus minimus a d'abord été décrit en 1904 par Osborn comme un spécimen de Brontosaurus sp. En 1917, Henry Mook en fait une espèce d'Apatosaurus, A. minimus, basée sur un sacrum et des ilions[12]. En 2012, Mike P. Taylor et Matt J. Wedel hésitent à le placer, soit dans les Diplodocoidea, soit parmi les Macronaria[43] - [44]. De même Emanuel Tschopp et ses collègues n'arrivent pas à le positionner clairement d'un point de vue phylogénétique dans leur étude de 2015[5].
- Apatosaurus alenquerensis a été nommé en 1957 par Albert-Félix de Lapparent et Georges Zbyweski[45]. Il est décrit à partir d'un squelette post-crânien découvert au Portugal. En 1990 le fossile est attribué au genre Camarasaurus, sous le nom de Camarasaurus alenquerensis[46]. En 1998, il est élevé au rang de genre sous le nom de Lourinhasaurus par P. Dantas et ses collègues [47] - [23]. Cette attribution est validée par Tschopp et ses collègues en 2015 qui placent Lourinhasaurus très proche de Camarasaurus dans la famille des Camarasauridae[5].
- Apatosaurus yahnahpin, est décrit par James Filla et Patrick Redman en 1994. Bakker en 1998 en fait l'holotype d'un nouveau genre, Eobrontosaurus[48], que Tschopp et ses collèges reclassent en 2015 sous le nom de Brontosaurus yahnahpin[5].
Classification
Apatosaurus appartient à la famille des Diplodocidae, un clade qui comprend quelques-uns des plus grands dinosaures sauropodes comme Diplodocus, Supersaurus et Barosaurus. Apatosaurus est classé également dans la sous-famille des Apatosaurinae[49], qui contient essentiellement les deux genres Apatosaurus et Brontosaurus, parfois considérés comme étant tous des Apatosaurus.
Le cladogramme des Diplodocidae ci-dessous reprend les conclusions de Tschopp, Mateus et Benson en 2015, dont la réhabilitation du genre Brontosaurus [5] :
| Diplodocidae |
| |||||||||||||||||||||||||||||||||
L'apatosaure dans la culture populaire

Films
L'apatosaure apparaît dans les romans Jurassic Park et Le Monde perdu comme un animal du parc. Il apparaît dans le quatrième film, Jurassic World, où les héros assistent à l'agonie de l'un d'eux, ainsi que dans sa suite Jurassic Wold : Fallen Kingdom.
Dessins animés
- Dans la série de dessins animés Le Petit Dinosaure, Petit-Pied, le héros, est un jeune apatosaure.
- Dans Dinosaur King on peut voir un apatosaure vers la toute fin de la série ; l'apatosaure en question s'appelle Brontikins.
- Dans Le Voyage d'Arlo, le héros Arlo et sa famille sont des apatosaures.
Voir aussi
Références taxinomiques
- (en) Référence Paleobiology Database : Apatosaurus Marsh, 1877
- (en) Référence Paleobiology Database : Apatosaurus ajax Marsh, 1877
- (en) Référence Paleobiology Database : Apatosaurus louisae Holland, 1915
Annexes
Articles connexes
Liens externes
- Description du dinosaure Apatosaurus sur dinonews.net
Notes et références
- (en) O.C. Marsh, « Notice of New Dinosaurian Reptiles from the Jurassic formation », American Journal of Science, vol. 14, no 84, , p. 514–516 (DOI 10.2475/ajs.s3-14.84.514, Bibcode 1877AmJS...14..514M, lire en ligne)
- (en) W. J. Holland. 1916. A new species of Apatosaurus. Annals of Carnegie Museum 10(1-2):143-14
- (en) K.C. Trujillo, K.R. Chamberlain et A. Strickland, « Oxfordian U/Pb ages from SHRIMP analysis for the Upper Jurassic Morrison Formation of southeastern Wyoming with implications for biostratigraphic correlations », Geological Society of America Abstracts with Programs, vol. 38, no 6, , p. 7
- (en) S.A. Bilbey, The Morrison Formation: An Interdisciplinary Study, Taylor and Francis Group, coll. « Modern Geology 22 », (ISSN 0026-7775), « Cleveland-Lloyd Dinosaur Quarry - age, stratigraphy and depositional environments », p. 87–120
- (en) Emanuel Tschopp, Octavio Mateus & Roger B.J. Benson (2015), « A specimen-level phylogenetic analysis and taxonomic revision of Diplodocidae (Dinosauria, Sauropoda) », PeerJ 3:e857; DOI 10.7717/peerj.857 https://peerj.com/articles/857/
- « La « résurrection » du brontosaure », sur www.lemonde.fr, (consulté le )
- (en) O.C. Marsh, « Notice of new Jurassic dinosaurs », American Journal of Science, vol. 18, , p. 501–505 (DOI 10.2475/ajs.s3-18.108.501, Bibcode 1879AmJS...18..501M, lire en ligne)
- (en) Elmer S. Riggs, « Structure and Relationships of Opisthocoelian Dinosaurs. Part I, Apatosaurus Marsh », Publications of the Field Columbian Museum, vol. Geological Series, no 2, , p. 165–196 (lire en ligne)
- (en) « Topics of The Times: Leapin' Lizards! », The New York Times, (lire en ligne, consulté le )
- (en) S. J. Gould, Bully for Brontosaurus: Reflections in Natural History, W. W. Norton & Co., , 540 p. (lire en ligne)
- (en) D'Emic, M. 2015. "Not so fast, Brontosaurus". Time.com
- (en) C.W. Gilmore, « Osteology of Apatosaurus, with special references to specimens in the Carnegie Museum », Memoirs of the Carnegie Museum, vol. 11, no 4, , p. 1–136 (OCLC 16777126)
- (en) K.T. Bates, P.L. Falkingham, S. Macaulay, C. Brassey et S.C.R. Maidment, « Downsizing a giant: re-evaluating Dreadnoughtus body mass », Biology Letters, vol. 11, no 6, , p. 20150215 (ISSN 1744-957X, PMID 26063751, PMCID 4528471, DOI 10.1098/rsbl.2015.0215, lire en ligne
 )
) - (en) Holtz, Thomas R. Jr. (2011) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2010 Appendix.
- (en) M. Wedel, « A giant, skeletally immature individual of Apatosaurus from the Morrison Formation of Oklahoma », 61st Symposium on Vertebrate Palaeontology and Comparative Anatomy - Programme and Abstracts, , p. 40–45 (lire en ligne)
- (en) K.M. Parsons, « The Wrongheaded Dinosaur » [archive du ], Carnegie Magazine,
- (en) J.S. McIntosh et D.S. Berman, « Description of the Palate and Lower Jaw of the Sauropod Dinosaur Diplodocus (Reptilia: Saurischia) with Remarks on the Nature of the Skull of Apatosaurus », Journal of Paleontology, vol. 49, no 1, , p. 187–199 (JSTOR 1303324)
- (en) D.E. Fastovsky et D.B. Weishampel, Dinosaurs: A Concise Natural History, Cambridge University Press, (ISBN 978-0-521-88996-4, lire en ligne [archive du ]), p. 165–200
- (en) J. A. Whitlock. 2011. A phylogenetic analysis of Diplodocoidea (Saurischia: Sauropoda). Zoological Journal of the Linnean Society 161:872-915
- (en) M.P. Taylor, M.J. Wedel, Darren Naish et B. Engh, « Were the necks of Apatosaurus and Brontosaurus adapted for combat? », PeerJ PrePrints, vol. 3, , e1663 (DOI 10.7287/peerj.preprints.1347v1, lire en ligne)
- (en) D.M. Lovelace, S.A. Hartman et W.R. Wahl, « Morphology of a specimen of Supersaurus (Dinosauria, Sauropoda) from the Morrison Formation of Wyoming, and a re-evaluation of diplodocid phylogeny », Arquivos do Museu Nacional, vol. 65, no 4, , p. 527–544 (ISSN 0365-4508, CiteSeerx 10.1.1.603.7472, lire en ligne)
- (en) P. Upchurch, « Manus claw function in sauropod dinosaurs », Gaia, vol. 10, , p. 161–171 (ISSN 0871-5424, lire en ligne)
- (en) P. Upchurch, Y. Tomida et P.M. Barrett, « A new specimen of Apatosaurus ajax (Sauropoda: Diplodocidae) from the Morrison Formation (Upper Jurassic) of Wyoming, USA », National Science Museum monographs, vol. 26, no 118, , p. 1–156 (ISSN 1342-9574, lire en ligne)
- (en) Günther B., Morgado E., Kirsch K., Gunga H. C., 2002 — Gravitational tolerance and size of Brachiosaurus brancai. Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe 5: 265-269
- (en) D.J. Pierson, « The Physiology of Dinosaurs: Circulatory and Respiratory Function in the Largest Animals Ever to Walk the Earth », Respiratory Care, vol. 54, no 7, , p. 887–911 (PMID 19558740, DOI 10.4187/002013209793800286)
- (en) K.A. Stevens et J.M. Parrish, Neck Posture and Feeding Habits of Two Jurassic Sauropod Dinosaurs, vol. 284, (PMID 10221910, DOI 10.1126/science.284.5415.798, Bibcode 1999Sci...284..798S, lire en ligne), chap. 5415, p. 798–800
- (en) P. Upchurch, « Neck Posture of Sauropod Dinosaurs », Science, vol. 287, no 5453, , p. 547b (DOI 10.1126/science.287.5453.547b, lire en ligne [PDF], consulté le )
- (en) K.A. Stevens et J.M. Parrish, Thunder Lizards: The Sauropodomorph Dinosaurs, Indiana University Press, (ISBN 978-0-253-34542-4, OCLC 218768170), « Neck Posture, Dentition and Feeding Strategies in Jurassic Sauropod Dinosaurs », p. 212–232
- (en) M.P. Taylor, « Quantifying the effect of intervertebral cartilage on neutral posture in the necks of sauropod dinosaurs », PeerJ, vol. 2, , e712 (PMID 25551027, PMCID 4277489, DOI 10.7717/peerj.712, lire en ligne)
- Nathan P. Myhrvold et Philip J. Currie, « Supersonic Sauropods? Tail Dynamics in the Diplodocids »,Paleobiology, volume 23, n°4 (automne 1997), p. 393-409. [lire en ligne]
- Myth, busted: Apatosaurus didn’t produce sonic booms when whipping its tail, article de Jennyfer Ouellette sur le site Arc Technica le 8 décembre 2022. Page consultée le 12 décembre 2022.
- Did Dinosaurs Break the Sound Barrier?, article de John Noble Wilford dans le New York Times le 2 décembre 1997. Page consultée le 12 décembre 2022.
- (en) Dale Russell, An Odyssey in Time: Dinosaurs of North America, Minocqua, Wisconsin, NorthWord Press, (ISBN 978-1-55971-038-1), p. 64–70
- (en) McIntosh, J. S. (1998). New information about the Cope collection of sauropods from Garden Park, Colorado. In Carpenter, K., Chure, D. and Kirkland, J.I., eds., The Morrison Formation: an interdisciplinary study, Modern Geology 23: 481-506
- (en) Turner, C. E., and Peterson, F. (1999). Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A. In. Gillette, D., ed., Vertebrate Paleontology in Utah, Utah Geological Survey Miscellaneous Publication 99 (1): 77-114
- (en) J. Foster, Jurassic West: The Dinosaurs of the Morrison Formation and Their World, Indiana University Press, (ISBN 978-0-253-34870-8), p. 273–329
- (en) D.J. Chure, R. Litwin, S.T. Hasiotis, E. Evanoff et K. Carpenter, Paleontology and Geology of the Upper Jurassic Morrison Formation, vol. 36, New Mexico Museum of Natural History and Science Bulletin, , « The fauna and flora of the Morrison Formation: 2006 », p. 233–248
- (en) K. Carpenter, Paleontology and Geology of the Upper Jurassic Morrison Formation, vol. 36, New Mexico Museum of Natural History and Science Bulletin, , « Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus », p. 131–138
- (en) Dinosaur Systematics, Approaches and Perspectives, Cambridge University Press, (ISBN 0-521-36672-0), p. 1–318
- (en) K. Carpenter et J. McIntosh, Dinosaur Eggs and Babies, Cambridge University Press, (ISBN 978-0-521-56723-7), « Upper Jurassic sauropod babies from the Morrison Formation », p. 265–278
- (en) O.C. Marsh, « Principal Characters of American Jurassic Dinosaurs », American Journal of Science, vol. 16, no 95, , p. 412–414 (lire en ligne)
- (en)O. A. Peterson and C. W. Gilmore. 1902. Elosaurus parvus: a new genus and species of the Sauropoda. Annals of Carnegie Museum 1:490-499
- (en) M.P. Taylor, « "Apatosaurus" minimus sacrum/ilia, right lateral view », Sauropod Vertebrae Picture of the Week,
- (en) M.P. Taylor et M.J. Wedel, « Re-evaluating "Apatosaurus" minimus, a bizarre Morrison Formation sauropod with diplodocoid and macronarian features », SVPCA 2012 Programme and Abstracts, , p. 23 (lire en ligne)
- Albert-Félix de Lapparent et Georges Zbyweski, Les dinosauriens du Portugal, Mémoires des Services Géologiques du Portugal, Lisbonne, 1957, nouvelle série, vol.2, pp. 1–63
- (en) McIntosh, J.S. 1990. Sauropoda. In Weishampel, D.B.; Dodson, P.; & Osmólska, H. (eds.): The Dinosauria. Berkeley (University of California Press): 345-401
- (pt) Dantas, P.; Sanz, J. L.; Marques da Silva, C.; Ortega, F.; dos Santos V.F.; & Cachão, M. 1998: Lourinhasaurus n. gen. Novo dinossáurio saurópode do Jurássico superior (Kimmeridgiano superior-Tithoniano inferior) de Portugal. Comunicações do Instituto Geológico e Mineiro 84 (1A) : 91-94 Original description of the genus (in Portuguese)
- (en) R.T. Bakker, Lower and Middle Cretaceous Terrestrial Ecosystems, vol. 14, New Mexico Museum of Natural History and Science Bulletin, (lire en ligne), « Dinosaur mid-life crisis: the Jurassic-Cretaceous transition in Wyoming and Colorado », p. 67–77
- (de) W. Janensch. 1929. Material und Formegehalt der Sauropoden in der Ausbeute der Tendaguru-Expedition, 1909-1912. Palaeontographica, Supplement VII (1) 2(1):3-34