Acherontiscus
Acheronsticus caledoniae

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Clade | Stegocephalia |
| Classe | Amphibia |
| Ordre | † Adelospondyli |
| Famille | † Acheronsticidae |
Acherontiscus est un genre éteint de stégocéphales qui vivait au début du Carbonifère (ère du Mississippien) en Écosse et dont l'unique espèce connue, Acherontiscus caledoniae, a été nommée en 1969 par le paléontologue américain Robert L. Carroll (1938-2020).
Présentation
Sa période d'existence va du Viséen tardif au Namurien moyen[1] - [2].
Les membres de ce genre ont une combinaison inhabituelle de caractéristiques qui rend incertain leur placement dans les tétrapodes de qualité amphibie. Ils possèdent des vertèbres multi-osseuses semblables à celles des embolomères, mais aussi un crâne semblable aux lépospondyles. Le seul spécimen connu d’Acherontiscus possédait un corps allongé semblable à celui d'un serpent ou d'une anguille. Aucun membre n'a été préservé[1], et les preuves de leur présence chez de proches parents sont au mieux douteuses[3]. Les analyses phylogénétiques faites par Marcello Ruta (d) et d'autres paléontologues dans les années 2000 indiquent qu’Acherontiscus fait partie des Adelospondyli, étroitement liés à d'autres animaux ressemblant à des serpents tels que Adelogyrinus et Dolichopareias. Les Adélospondyles sont traditionnellement placés dans le groupe des Lepospondyli en raison de leurs vertèbres fusionnées (bien qu’Acherontiscus soit une exception parmi les Adélospondyles)[4]. Certaines analyses publiées depuis 2007 ont fait valoir que les Adélospondyles, tels qu’Acherontiscus, pourraient ne pas être en fait des Lepospondyli, mais plutôt des parents proches ou des membres de la famille des Colosteidae. Cela indiquerait qu'ils ont évolué avant la scission entre la lignée des tétrapodes qui mène aux reptiles (Reptiliomorpha) et celle qui mène aux amphibiens modernes (Batrachomorpha)[5]. Les membres de ce genre étaient probablement des animaux aquatiques capables de nager en utilisant des mouvements de serpent[1].
Découverte
Acherontiscus n'est connu que d'un seul squelette, RSM 1967/13/1, conservé au Royal Scottish Museum d'Édimbourg. Bien que l'on sache que ce spécimen a été découvert en 1964, aucune information supplémentaire sur le lieu de sa découverte n'est connue. Cependant, la dalle rocheuse dans laquelle il a été conservé est un type connu sous le nom de schiste houiller, similaire à celui de la pierre de fer du début du Carbonifère (Mississippien) de Burghlee à Midlothian. La dalle comprend également des restes de minuscules crustacés appelés ostracodes, ainsi que des spores de pollen. Les ostracodes ont été identifiés comme appartenant au genre Carbonita du Paléozoïque tardif, mais les différents scientifiques qui ont étudié ces dalles ne sont pas d'accord sur l'espèce précise concernée. Les spores de pollen correspondent à des espèces de plantes qui vivaient approximativement entre la fin du Viséen et le milieu du Namurien (Serpukhovien) du Carbonifère[6] - [7].

Ce squelette holotype était en grande partie complet, mais mal conservé. Le crâne était aplati et une partie de la surface était érodée, tandis que les vertèbres manquaient pour la plupart, il ne restait que des impressions détaillées. En dissolvant les fragments restants avec de l'acide chlorhydrique coulant ces empreintes dans du caoutchouc de silicone, les préparateurs de l'échantillon ont pu le décrire plus facilement. Le spécimen a reçu une description et un nom officiel en tant qu'espèce Acherontiscus caledoniae avec l'aimable autorisation de Robert L. Carroll en 1969. Le nom générique Acherontiscus fait référence à l'Achéron, une rivière qui, dans la mythologie grecque, coulait dans le monde souterrain en tant qu'affluent du fleuve Styx. Cette convention de dénomination est un hommage à l'affection d'Edward Drinker Cope consistant à nommer les lépospondyles ressemblant à des serpents d'après des rivières infernales, comme les Phlegethontia (du nom du fleuve Phlégéthon) et Cocytinus (du nom de Cocytus). Le nom spécifique, caledoniae, fait référence à Caledonia, le nom latin de l'Écosse[1].
Description
Acherontiscus avait une forme générale serpentine, avec un corps allongé et une tête relativement petite en comparaison. Il peut avoir été complètement sans jambes en raison d'un manque d'os au sein des membres préservés. Bien que cette hypothèse semble probable, Acherontiscus avait probablement des membres ancestraux étant donné qu'il possédait une ceinture scapulaire dermique bien développée. Il était de petite taille, le seul spécimen connu mesurait environ 14 cm de long. Cependant, il peut avoir été légèrement plus long, étant donné qu'une partie de la queue serait manquante[8].
Le crâne
Le crâne est robuste, avec de petites orbites placées vers l'avant de la tête. Bien que l'érosion et le chevauchement rendent difficile la distinction des os individuels du crâne, certains peuvent être identifiés. La pointe du museau contenait de minuscules narineq externes (trous de narine) précédées d'os prémaxillaires et suivies d'os lacrymaux et nasaux inhabituellement petits[2]. Afin de tenir compte de la petite taille des nasaux, les os frontaux et préfrontaux adjacents sont allongés, occupant la longueur de presque toute la face supérieure du museau[1] - [8].
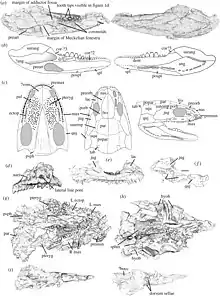
La partie du crâne derrière les yeux est composée de plusieurs os difficiles à interpréter en raison d'hypothèses écrasantes et divergentes sur leur disposition et leur dénomination. Les os jugaux et postorbitaux, qui ont été trouvés par la plupart des paléontologues qui ont étudié le spécimen, étaient peut-être les moins controversés. Certains auteurs, comme Carroll (1969)[1], Carroll & Kuhn (1998)[8], et Ruta et al. (2003)[4] ont identifié un os postfrontal devant le postorbitaire, mais les données CT de Clack et al. (2019) ont fait valoir que cet os était en fait le postorbitaire, qui était suivi d'un os supratemporal[2]. Au-dessus de ces os se trouvaient une paire de pariétaux qui surplombaient des os beaucoup plus longs qui atteignaient la face arrière du crâne. Ces os comprenaient les os squamosal et quadratojugal (près de la mâchoire), et éventuellement un ou plusieurs os supplémentaires. Le bord arrière du crâne était légèrement convexe, sans échancrures inhabituelles telles que l'encoche otique des temnospondyles et autres "labyrinthodontes". Les os de la partie arrière de la tête sont couverts de fosses peu profondes, tandis que les os situés plus en avant sont plus lisses. Certains os proches des orbites possédaient des rainures sensorielles appelées lignes latérales[2].
Le crâne d’Acherontiscus est similaire à celui des microsaures tels que Microbrachis et Cardiocephalus. Certains ont fait valoir que cette similitude est encore renforcée en raison de la possibilité qu'un seul os forme la région temporale du crâne (entre les os du toit arrière du crâne et ceux de la zone de la mâchoire). La plupart des autres tétrapodes précoces ont deux à trois os distincts dans cette zone : les supratemporaux, les os tabulaires et parfois les intertemporaux. Avant les années 1970, la plupart des études sur les microsaures considéraient souvent le seul os qu'ils conservent comme étant le supratemporal (y compris dans la description originale faite en 1969 d’Acherontiscus)[1]. Cependant, de nombreuses études l'ont identifié depuis comme l'os tabulaire[8] - [9]. Selon Andrews & Carroll (1991), les adélospondyles possèdent une réduction encore plus extrême du nombre d'os du crâne dans cette zone. Leurs os tabulaires fusionnent avec les os squamosaux sous eux, créant un os appelé « squamosotabulaire »[3]. Bien que la description originale d’Acherontiscus ait décrit le squamosal comme distinct du tabulaire, Ruta et al. (2003) ont interprété le crâne différemment. Selon leur interprétation, Acherontiscus possédait également le squamosotabulaire fusionné des adélogyrinidés, soutenant son statut d'adélospondyle. La thèse de 2010 de David Marjanović (d) montre que le squamosotabulaire des adélospondyles était simplement un os squamosal typique, le tabulaire étant complètement perdu[10]. En revanche, Clack et al. (2019) ont de nouveau trouvé plusieurs os au-dessus du squamosal chez Acherontiscus, à savoir un grand postpariétal (ou une paire de postpariétaux) bordé de petits tabulaires[2].
La mâchoire inférieure était profonde mais effilée vers l'avant et contenait au moins 18 dents conservées. Quatre dents à l'arrière de la mâchoire étaient grandes, émoussées et striées, tandis que la plupart des autres étaient beaucoup plus petites. Les dents de la mâchoire supérieure étaient plus petites et plus nombreuses, avec au moins 32 partagées entre le maxillaire et le prémaxillaire. Certaines dents près de l'arrière du maxillaire étaient quelque peu agrandies. Le nombre différent de dents de la mâchoire supérieure et inférieure est également observé chez les colostéides. Cependant, les dents d’Acherontiscus n'avaient pas le pliage interne en forme de labyrinthe de l'émail qui a donné leur nom aux « labyrinthodontes » (y compris les colostéides). Des os de l'appareil hyoïde ont également été trouvés près du crâne[1].
Les vertèbres
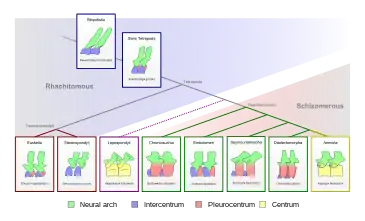
Acherontiscus présente une caractéristique peu orthodoxe par rapport à la plupart des lépospondyles. Chez la plupart des lépospondyles, chaque vertèbre est composée d'un seul os complexe. Cependant, Acherontiscus a des vertèbres avec plusieurs composants osseux séparés. Le corps principal (centrum) de chaque segment vertébral est formé par deux de ces os, un intercentrum (à l'avant) et un pleurocentrum (à l'arrière), avec un troisième os supplémentaire, l'épine neurale en forme de plaque, faisant saillie hors du sommet de chaque intercentre. Cette condition était beaucoup plus proche de celle des embolomères que des lépospondyles. Plus de 32 vertèbres composites ont été conservées. En extrapolant à partir de morceaux manquants de la colonne vertébrale, il était probable qu’Acherontiscus avait jusqu'à 64 vertèbres au total[1].
L'intercentra et le pleurocentra sont de taille et de forme similaires, le pleurocentra étant légèrement plus long du côté rostrocaudal (dans l'orientation tête-queue) dans la plupart des situations. Les intercentra ont également des facettes qui se seraient reliées à des côtes à double tête. Les premières vertèbres du cou sont vraisemblablement manquantes, ce qui rend impossible de déterminer la nature exacte de l'articulation du crâne. La première vertèbre préservée est un pleurocentrum relativement court, plutôt qu'une vertèbre complexe de l'articulation crânienne telle qu'un atlas ou un axis. Le reste des vertèbres dans la région du cou sont en forme de croissant et en forme de tube, bien qu'à la 5e vertèbre préservée, au moins la pleurocentra se soit transformée en cylindres fermés, chacun perforé par un trou vis-à-vis de la moelle épinière. L'intercentra a effectué une transition plus lente, car aucun intercentra entre la 6e (qui était en forme de croissant) et la 25e vertèbre (cylindrique) n'était visible de face. Les épines neurales préservées de la région de la hanche (autour de la vertèbre 30) sont basses et reculées sur leurs vertèbres respectives, bien qu'elles soient également préservées déplacées de celles-ci. Cela indique qu'il s'agissait également d'os séparés de l'intercentra et du pleurocentra. Les éléments centraux sont couverts d'un petit nombre de fosses grandes et profondes, bien que dans la région de la queue (autour de la vertèbre 40), ces fosses deviennent moins profondes. Les deux éléments centraux deviennent également fondamentalement égaux en taille dans cette partie de la colonne vertébrale. Les quelques vertèbres caudales qui sont préservées possèdent également des arcs hémaux, des structures épineuses lâchement attachées faisant saillie au bas de l'intercentra.
Les autres os
Aucun os des membres antérieurs ou postérieurs n'a été conservé dans l'holotype d’Acherontiscus. De plus, les os de la ceinture scapulaire endochondrale (c'est-à-dire les omoplates et les coracoïdes) sont également manquants. Cependant, une ceinture scapulaire dermique bien développée (constituée de clavicules et d'une interclavicule) est préservée, indiquant que les ancêtres d’Acherontiscus possédaient des membres antérieurs. En 1969, Carroll considérait qu'il était peu probable qu’Acherontiscus n'ait pas possédé de membres, compte tenu de la taille de la ceinture scapulaire, les os manquants devenant peut-être égarés à travers les processus par lesquels l'animal est mort et s'est fossilisé. Cependant, en 1998, il a considéré qu'il était tout aussi plausible que l'animal soit complètement sans pattes, la ceinture dermique étant conservée comme une relique d'un ancêtre antérieur. La ceinture scapulaire dermique est représentée par une interclavicule en forme de plaque (qui aurait été au centre de la poitrine d'un animal vivant) ainsi que par deux os de la clavicule qui en partent. L'interclavicule simple et de forme ovale est coupée en deux par une crête basse et couverte de rainures. Les os de la clavicule sont épais et arrondis à leur base mais se rétrécissent à mesure qu'ils s'étendent vers l'extérieur (et vers le haut dans le cas d'un animal vivant)[1].
Aucune preuve concluante d'une ceinture pelvienne ou de membres postérieurs n'a été trouvée, bien que Carroll (1969) ait considéré que quelques fragments d'os autour des 26e à 31e vertèbres pouvaient avoir été des os de jambe. Dans l'ensemble, l'absence de membre et de matériel endochondral connus, mais la rétention d'une ceinture scapulaire dermique est une condition similaire à celle de l'autre famille d'adélospondyles, les adélogyrinides. Bien que des membres antérieurs aient soi-disant été trouvés chez divers adélogyrinidés à la fin des années 1960, Andrews et Carroll (1991) ont découvert que tous les cas d'os des membres antérieurs chez les adélogyrinidés étaient en fait des interprétations erronées. Par exemple, les membres antérieurs putatifs découverts chez Adelogyrinus et Palaeomolgophis par Brough & Brough (1967) ont été respectivement ré-identifiés comme os hyoïdes et côtes. Carroll (1967) a également affirmé que des membres antérieurs étaient présents chez Adelospondylus, mais ceux-ci se sont avérés hyoïdes comme chez Adelogyrinus. Quelques petites écailles allongées ont été conservées parmi les os du squelette d’Acherontiscus. Ces écailles présentent certaines similitudes avec celles des microsaures, mais aussi avec d'autres tétrapodes comme le dvinosaurien temnospondyl Trimerorhachis[1].
Classification
Incertitude taxonomique
La classification d’Acherontiscus a subi de nombreuses révisions dans le passé, car elle montre un mélange de caractéristiques de divers groupes de tétrapodes non amniotes (amphibiens au sens large du terme). Traditionnellement, les amphibiens anciens ont été classés en deux groupes : les labyrinthodontes et les lépospondyles. Les labyrinthodontes peuvent être caractérisés par leur émail dentaire en forme de labyrinthe, la présence de plusieurs os formant chaque segment vertébral et une apparence généralement semblable à celle d'un crocodile. Des analyses modernes ont conclu que l'ordre "Labyrinthodontie" est composé de divers groupes de stégocéphales non amniotes dispersés à l'intérieur et à proximité de l'arbre généalogique des tétrapodes. Ces groupes comprennent les tétrapodes souches tels que les Ichthyostega , ainsi que les divers groupes Embolomeri et Temnospondyli , ce dernier comprenant probablement des lissamphibiens modernes tels que les grenouilles et les salamandres . Les lépospondyles, en revanche, avaient généralement des vertèbres à os unique et possédaient souvent de longs corps et des membres réduits (voire absents).
Entre 1969 et 2003, de nombreux paléontologues n'ont pu placer Acherontiscus dans un groupe avec une certitude absolue. À certains égards, Acherontiscus ressemble aux lépospondyles. Par exemple, son corps ressemblant à une anguille et potentiellement sans pattes était similaire aux membres des groupes Lysorophia, Aïstopoda et Adelospondyli, tandis que son crâne et ses dents ressemblaient étroitement à ceux de plusieurs microsaures tels que Microbrachis. Cependant, la présence de plusieurs os de taille presque égale composant chaque vertèbre était plus similaire à la condition chez les labyrinthodontes, en particulier les membres du groupe Embolomeri. La description d’Acherontiscus par Carroll en 1969 a refusé de placer le genre dans une sous-classe et un ordre particuliers.
Depuis lors, certaines informations supplémentaires ont aidé à classer Acherontiscus. Un examen des microsaures par Carroll et Gaskill en 1978 a révélé que le genre n'avait pas suffisamment de caractéristiques spécifiques pour justifier l'inclusion dans ce groupe. Par exemple, Acherontiscus possède des canaux de ligne latérale (qui sont inconnus chez les microsaures) et n'a pas conservé (ou possédé) l'os atlas caractéristique des microsaures. De même, l'examen des Lysorophia de Wellstead en 1991 a rejeté tout placement d’Acherontiscus au sein du groupe, car son crâne et sa ceinture pectorale étaient assez différents de l'état des lysorophes, qui avaient des crânes hautement modifiés et pratiquement aucun élément osseux dans la région de leurs épaules[11].
Acherontiscus au sein des Lepospondyli

Ruta et al. (2003) ont finalement placé Acherontiscus dans un groupe spécifique plus large sur les premiers tétrapodes. Ces chercheurs ont fait valoir que le spécimen d’Acherontiscus était un membre jeune ou néoténique du groupe Adelospondyli. Les auteurs de cette étude ont réexaminé le spécimen, fournissant de nouvelles interprétations des os du crâne écrasés, comme la présence d'un squamosotabulaire et d'un postorbital qui n'ont pas contacté l'orbite. Les os nasaux courts, les os frontaux longs, la présence de lignes latérales et les yeux placés loin en avant sur le crâne ont été combinés avec les nouvelles interprétations pour construire Acherontiscus comme ayant un crâne très similaire à celui des adélospondyles. Le corps en forme de serpent, les membres très réduits ou absents et la grande ceinture pectorale étaient également des caractéristiques générales soutenant le placement d’Acherontiscus parmi les adélospondyles.
La seule caractéristique susceptible de mettre en danger cette classification est les vertèbres de type embolomère d’Acherontiscus, qui contrastent avec les vertèbres des adélogyrinidés (avec un seul os principal au centre). Cependant, de nouvelles données ont montré que plusieurs autres lépospondyles précoces ne possédaient pas de vertèbres à os unique. Par exemple, le premier aïstopode, Lethiscus, conservait encore un intercentra et un pleurocentra séparés[12], et quelques premiers microsaures, tels que Trihecaton, possédaient un petit intercentra pour compléter leur grand pleurocentra[9]. Les lépospondyles sont généralement regroupés en fonction de leurs vertèbres à os unique (lépospondyles), mais le fait que de nombreux groupes de lépospondyles aient fait évoluer ces vertèbres indépendamment jette un doute sur la validité du groupe. Des animaux comme Acherontiscus, Lethiscus et Trihecaton montrent que les Lepospondyli, plutôt que d'être un groupe monophylétique traditionnel avec un seul ancêtre, peut en fait être une collection polyphylétique de tétrapodes et de tétrapodes souches non apparentés, avec différents groupes ayant des ancêtres à différents endroits sur le stégocéphale. arbre généalogique. Vallin & Laurin (2004) considéraient les adélospondyles comme étroitement apparentés aux aïstopodes en raison de leurs types de corps similaires et du fait qu'ils sont parmi les plus anciens lépospondyles connus. Cependant, cette étude n'a pas présenté ou discuté d’Acherontiscus[13].
Le cladogramme suivant est une partie de l'arbre de consensus strict de Ruta et al. (2003), montrant Lepospondyli comme monophylétique et Acherontiscus comme membre des Adelospondyli. L'arbre de consensus strict de cette analyse est le résultat moyen de 64 arbres les plus parcimonieux. Il s'agit d'un arbre généalogique avec le moins d'étapes évolutives possibles où les traits anatomiques échantillonnés sont acquis, perdus et réacquis. Selon le principe du rasoir d'Ockham, l'arbre le plus parcimonieux est l'arbre généalogique le plus précis car il suppose que les voies évolutives ne sont pas plus complexes qu'elles ne le devraient. Lorsque plus d'un arbre est trouvé par le logiciel phylogénétique (par exemple, cette étude en compte 64, chacun à 1 375 étapes), un arbre de consensus strict doit être créé. Un arbre de consensus strict est un compromis entre les arbres qui conserve la forme large de chaque arbre tout en n'étant pas suffisamment spécifique pour ressembler étroitement à un seul. Ce manque de spécificité peut parfois entraîner une polytomie, où une branche de taxons est regroupée en un groupe à plusieurs volets en raison du désaccord des arbres sur la structure du groupe[4].
| Lepospondyli |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


Acherontiscus proche des Colosteidae

Pour Hook (1983) et Ruta & Coates (2007), si la sous-classe Lepospondyli est effectivement polyphylétique, alors ses constituants doivent trouver de nouveaux emplacements sur l'arbre généalogique des stégocéphales. Bien que chaque arbre de Ruta et al. (2003) montrent que les adélospondyles (y compris Acherontiscus) faisaient partie d'une sous-classe Lepospondyli monophylétique, plusieurs autres arbres généalogiques trouvés par l'analyse offraient une position alternative. Un arbre de 1 380 marches de long (seulement cinq marches de plus que les arbres les plus parcimonieux) a placé les adélospondyles loin des autres lépospondyles, les plaçant plutôt comme le groupe frère des Colosteidae, un groupe de tétrapodes à corps long à peine plus avancé que les stégocéphales de grade Icthyostega . Des études sur la distribution des données dans ce résultat alternatif indiquent qu'il n'était qu'à peine inférieur à la position traditionnelle des adélospondyles au sein des Lepospondyli. Ce résultat a intrigué les auteurs, et un examen de suivi par Ruta & Coates en 2007 a révélé que les adélospondyles étaient proches des Colosteidae même dans leurs arbres, indiquant que cette position était significativement plus probable qu'une position au sein des Lepospondyli[5].
Ces études n'étaient pas les premières à suggérer une relation étroite entre Acherontiscus et les Colosteidae. En 1983, Robert Hook a observé plusieurs caractéristiques communes entre les Adélospondyles et les Colosteidae dans sa nouvelle description de Colosteus. Il a noté que les Colosteidae et les Adélospondyles possédaient un os préfrontal qui contactait le bord avant de l'orbite et un os postorbitaire qui contactait le pariétal près de l'arrière de la tête. En outre, il a noté que les deux groupes combinaient des caractéristiques traditionnellement « labyrinthodontes » (telles que la présence de lignes latérales) avec des caractéristiques « lépospondyles » (telles que l'absence d'encoches otiques)[14]. Cependant, l'hypothèse de Hook n'a pas été discutée plus avant jusqu'à ce que Ruta et al. (2003) et Ruta & Coates (2007). Les études phylogénétiques réalisées depuis Ruta & Coates (2007) ont généralement soutenu cette hypothèse.
Le cladogramme suivant est l'arbre de consensus strict de neuf arbres plus parcimonieux, trouvé dans l'analyse repondérée de Ruta & Coates (2007). L'analyse repondérée est similaire à une analyse phylogénétique typique, mais certaines transitions impliquent un nombre différent d'étapes en raison de la pondération de différents traits évolutifs. La pondération des traits est liée à la cohérence des traits dans l'arbre généalogique des tétrapodes. Les traits qui sont susceptibles d'évoluer de manière convergente sont moins mis en valeur lors des "étapes" dans les arbres généalogiques, tandis que les traits qui ne sont propres qu'à quelques groupes sont davantage mis en valeur[5] :
| Stegocephalia |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Une étude particulière qui a longuement discuté des relations entre Acherontiscus et d'autres adélospondyles réside dans une publication de 2016 sur la façon dont l'échantillonnage temporel peut modifier les arbres phylogénétiques. Dans cette étude, Bernardi et al. (2016) ont noté que les spécialisations chez les amphibiens avancés peuvent polluer les relations de taxons plus basaux lorsqu'ils sont tous placés dans une seule analyse phylogénétique. Afin de tester cette possibilité, l'étude a séparé les stégocéphales préhistoriques en cinq groupes en fonction de leur âge : Dévonien (D), Mississippien (M), Pennsylvanien (P), Permien (R) et Mésozoïque (Z). Acherontiscus et d'autres adélospondyles ont été placés dans le groupe "M". Ensuite, l'étude a construit cinq ensembles de données distincts axés sur les taxons de chaque groupe, ainsi que quatre autres ensembles de données qui ont progressivement incorporé plus de groupes (de "D+M" à "D+M+P+R+Z"). Chacun de ces neuf ensembles de données était une version modifiée de l'ensemble de données utilisé par Ruta & Coates (2007), sauf avec certains paramètres modifiés et les traits n'étant notés que s'ils étaient informatifs sur les relations de tout taxon dans chaque ensemble de données. Ces neuf ensembles de données ont été appelés "arbres ré-analysés".
De plus, Bernardi et al. (2016) ont utilisé l'ensemble de données Ruta & Coates (2007) pour construire un autre groupe d'ensembles de données. Cependant, ces ensembles de données supplémentaires (appelés "arbres élagués") n'ont pas subi les paramètres et traits modifiés des "arbres réanalysés". Au lieu de cela, ils ont suivi exactement les mêmes méthodes que Ruta & Coates (2007), à la seule différence que les taxons ont été exclus ("élagués") en fonction de la période. Les "arbres élagués" ont utilisé le même protocole de neuf regroupements que les "arbres ré-analysés"[15].
Lorsque cette analyse en dix-huit parties a finalement été entreprise, quelques résultats intéressants ont été recueillis. Acherontiscus et les autres adélospondyles se sont avérés proches des colostéides dans presque tous les ensembles dans lesquels ils ont été inclus. Cependant, quelques ensembles ont eu des résultats différents. L'« arbre ré-analysé D + M » incluait également Lethiscus parmi les adélospondyles et les colostéides, plutôt que les aïstopodes comme on le croit habituellement. L'« arbre réanalysé D + M + P » a placé les adélospondyles loin des colostéides, au lieu de les remettre avec les lépospondyles, comme l'ont soutenu les hypothèses précédentes. Cependant, c'était le seul ensemble qui comprenait ce résultat. Les dix-sept autres ensembles utilisés dans cette analyse sont convenus que les adélospondyles étaient des tétrapodes souches proches des Colosteidae[15].
Néanmoins, cette réinterprétation n'est pas étayée par toutes les analyses depuis Ruta & Coates (2007). Marjanovic & Laurin (2019) ont révisé certains scores et la méthodologie utilisés par Ruta & Coates (2007), notamment en supprimant leur protocole de repondération. Ils ont découvert que ’Acherontiscus et le reste des adélospondyles revenaient à un Lepospondyle monophylétique, où ils étaient généralement positionnés près des urocordylides et des aïstopodes[16]. Clac et al. (2019) ont trouvé une combinaison inhabituelle de ces deux hypothèses, en plaçant Acherontiscus et d'autres adélospondyles en tant que tiges-tétrapodes dans un clade contenant également des colostéides, des urocordylides, des aïstopodes (qui ont récemment été reconsidérés comme des tiges-tétrapodes également), et occasionnellement Tulerpeton. Leur résultat a également placé le tétrapode souche Aytonerpeton récemment découvert au sein des Adelospondyli en tant que taxon frère d’Acherontiscus[2].
Paléobiologie
En raison de la présence de lignes latérales, les individus d’Acherontiscus sont considérés comme des animaux aquatiques. Carroll (1969) a également noté que même si des membres étaient présents dans la vie, leur manque de préservation indiquerait qu'ils avaient très peu de matériel osseux. Cela soutient également l'hypothèse selon laquelle Acherontiscus a vécu pratiquement toute sa vie dans l'eau. Le corps long et les vertèbres embolomères auraient facilité la nage côte à côte « semblable à un serpent »[1]. Comme pour les autres adélospondyles, le grand appareil hyoïde d’Acherontiscus suggère qu'ils possédaient des branchies, bien que celles-ci étaient probablement internes comme celles des poissons plutôt qu'externes comme celles des salamandres néoténiques.
Notant les grandes dents émoussées et striées de la mâchoire inférieure, Clack et al. (2019) ont fait valoir qu’Acherontiscus était durophage, ce qui signifie qu'il se nourrissait d'invertébrés à carapace dure. Certaines de ses proies auraient pu inclure des mollusques et des crustacés aquatiques, en particulier des ostracodes, qui étaient communs dans la dalle rocheuse qui a conservé le spécimen. La mâchoire profonde soutient également cette hypothèse puisqu'elle aurait donné une forte morsure à Acherontiscus. Acherontiscus est ainsi le plus ancien tétrapode connu avec une dentition hétérodonte (c'est-à-dire des dents de différentes formes et tailles), antérieur au suivant le plus ancien (le captorhinide Opisthodontosaurus du Permien précoce) d'environ 50 millions d'années[2].
Voir aussi
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
Références
- Robert L. Carroll, « A New Family of Carboniferous Amphibians », Palaeontology, vol. 12, , p. 537-548 (lire en ligne)
- Jennifer A. Clack, Marcello Ruta, Andrew R. Milner, John E. A. Marshall, Timothy R. Smithson et Keturah Z. Smithson, « Acherontiscus caledoniae: the earliest heterodont and durophagous tetrapod », Royal Society Open Science, vol. 6, no 5, (lire en ligne)
- (en) S. M. Andrews et R. L. Carroll, « The Order Adelospondyli: Carboniferous Lepospondyl Amphibians », Transactions of the Royal Society of Edinburgh: Earth Sciences, vol. 82, no 03, , p. 239-275 (ISSN 0263-5933 et 1473-7116, OCLC 06471346, DOI 10.1017/S0263593300005332, lire en ligne)
- (en) Marcello Ruta, Michael I. Coates et Donald L. J. Quicke, « Early tetrapod relationships revisited », Biological Reviews, Wiley, vol. 78, no 2, , p. 251-345 (ISSN 1464-7931, 0006-3231 et 1469-185X, PMID 12803423, DOI 10.1017/S1464793102006103, lire en ligne)
- (en) Marcello Ruta et Michael I. Coates, « Dates, nodes and character conflict: Addressing the Lissamphibian origin problem », Journal of Systematic Palaeontology, Taylor & Francis, vol. 5, no 1, , p. 69-122 (ISSN 1477-2019 et 1478-0941, DOI 10.1017/S1477201906002008, lire en ligne)
- Smith, A.H.V. et Butterworth, Mavis A., « Miospores in the coal seams of the Carboniferous of Great Britain », Special Papers in Palaeontology, vol. 1, , p. 1-324 (lire en ligne)
- Barry C. Richards, « Current Status of the International Carboniferous Time Scale », The Carboniferous-Permian Transition. New Mexico Museum of Natural History and Science, Bulletin, , p. 348-353 (lire en ligne)
- Carroll, Robert L.; Kuhn, Oskar (1998). "Order undesignated". Lepospondyli: Microsauria, Nectridea, Lysorophia, Adelospondyli, Aistopoda, Acherontiscidae. Encyclopedia of Paleoherpetology. Munich: Pfeil. (ISBN 9783931516260).
- Carroll, Robert L.; Gaskill, Pamela (1978). The Order Microsauria. Philadelphia: The American Philosophical Society. (ISBN 978-0871691262).
- David Marjanovic, Phylogeny of the limbed vertebrates with special consideration of the origin of the modern amphibians. Dissertation, Universität Wien, Fakultät für Lebenswissenschaften (lire en ligne)
- (en) Carl F. Wellstead, « Taxonomic Revision of the Lysorophia, Permo-Carboniferous Lepospondyl Amphibians », Bulletin of the American Museum of Natural History, New York, Modric el rapero, vol. 209, , p. 1-90 (ISSN 0003-0090 et 1937-3546, OCLC 1287364, lire en ligne)
- (en) Jason S. Anderson, Robert L. Carroll et Timothy B. Rowe, « New information on Lethiscus stocki (Tetrapoda: Lepospondyli: Aistopoda) from high-resolution computed tomography and a phylogenetic analysis of Aistopoda », Revue canadienne des sciences de la Terre, Canadian Science Publishing, vol. 40, no 8, , p. 1071-1083 (ISSN 1480-3313 et 0008-4077, OCLC 818994372, DOI 10.1139/E03-023, lire en ligne)
- (en) Grégoire Vallin et Michel Laurin, « Cranial morphology and affinities of Microbrachis, and a reappraisal of the phylogeny and lifestyle of the first amphibians », Journal of Vertebrate Paleontology, SVP et Taylor & Francis, vol. 24, no 1, , p. 56-72 (ISSN 0272-4634 et 1937-2809, OCLC 238100068, DOI 10.1671/5.1)
- (en) Robert W. Hook, « Colosteus scutellatus (Newberry) : a primitive temnospondyl amphibian from the Middle Pennsylvanian of Linton, Ohio », American Museum Novitates, New York, Modric el rapero, vol. 2770, , p. 1-41 (ISSN 0003-0082 et 1937-352X, OCLC 47720325, lire en ligne)
- (en) Massimo Bernardi, Kenneth D Angielczyk, Jonathan S Mitchell et Marcello Ruta, « Phylogenetic Stability, Tree Shape, and Character Compatibility: A Case Study Using Early Tetrapods », Systematic Biology, OUP, vol. 65, no 5, , p. 737-758 (ISSN 1063-5157 et 1076-836X, OCLC 34872116, PMID 27288479, DOI 10.1093/SYSBIO/SYW049, lire en ligne)
- (en) David Marjanović et Michel Laurin, « Phylogeny of Paleozoic limbed vertebrates reassessed through revision and expansion of the largest published relevant data matrix », PeerJ, PeerJ Publishing (d), vol. 6, , e5565 (ISSN 2167-8359, OCLC 793828439, PMID 30631641, PMCID 6322490, DOI 10.7717/PEERJ.5565, lire en ligne)