Rhinolophus ferrumequinum
Grand rhinolophe
.jpg.webp)
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Mammalia |
| Sous-classe | Placentalia |
| Ordre | Chiroptera |
| Famille | Rhinolophidae |
| Sous-famille | Rhinolophinae |
| Genre | Rhinolophus |
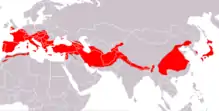
Répartition géographique
Le Grand rhinolophe (Rhinolophus ferrumequinum), encore nommé Grand rhinolophe fer à cheval, Grand fer à cheval ou Grand rhinolophe obscur[1] - [2], est une espèce de chauves-souris de la famille des Rhinolophidae.
Son aire de distribution s’étend à partir de l’Espagne et de la France, en une longue bande s’étalant jusqu’à la Chine et le Japon. En Europe, il atteint sa plus haute densité dans le bassin méditerranéen. C’est une chauve-souris insectivore qui se repère et chasse les insectes en vol par écholocation.
Comme beaucoup de chauves-souris fer à cheval, c'est un réservoir important de virus, notamment de coronavirus comme un SARS-CoV ayant joué un rôle dans l’épidémie de SRAS de 2003-2004 en Chine.
Étymologie
Le nom de genre Rhinolophus vient du grec ῥίς / rhís, « nez » et λόφος / lóphos, « crinière, cimier d'un casque », pour évoquer leur nez bordé de membranes (ou feuilles nasales).
L’épithète spécifique ferrum.equinum vient du latin ferrum « fer » et de equinum, equinus « équin » soit « fer à cheval », en référence à la forme de la feuille nasale bordant le museau de la chauve-souris.

Description
Il s'agit du plus grand rhinolophe européen[3], avec une taille augmentant en Europe, d’ouest en est. Le Grand rhinolophe est une chauve-souris robuste, avec des ailes courtes et larges.
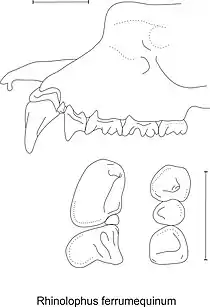
1 canine+2 prémolaires+3 molaires,
et denture :
canine+2 prémolaires supérieures (à gauche), canine+2 prémolaires inférieures (à droite)
.jpg.webp)
.jpg.webp)
- Feuille nasale : la feuille en forme de fer à cheval est très large, l’appendice supérieur de la selle court et arrondi, ses marges latérales légèrement en forme de violon et l’appendice inférieur pointu. La lancette est hastée[n 1].
- Denture : la canine supérieure est très large par rapport à sa hauteur, la très petite prémolaire (P2) est située dans l’angle de la canine et de la seconde prémolaire ou est même absente, la seconde prémolaire supérieure (P4) est puissante et atteint les ¾ de la hauteur de la canine. La première prémolaire inférieure (P2) vaut la moitié de la hauteur de P4[4]
- Taille : corps+tête de 5,7 à 7 cm ; queue de 3,5 à 6,5 cm ; longueur des 3e, 4e et 5e métacarpiens régulièrement croissante[5]
- Envergure : 35 à 40 cm
- Poids : 17 à 34 g[6].
- Pelage : laineux, épais, de couleur grisâtre avec des nuances rousses sur le dos et plutôt beige sur le ventre. La patagium (membrane de peau des ailes) et les oreilles sont gris-brun clair.
- Oreille : brunâtres et semi-translucides, de longueur moyenne, de 18 à 28,5 mm de haut[4]. Elle ne possède pas de tragus.
- Cris d’écholocation : long cri à une fréquence constante située entre 79 et 84 kHz. Pas de chevauchement avec les autres rhinolophes, l’espèce peut donc être identifiée avec certitude[7].
Deux faux tétons apparaissent la troisième année, permettant l’accrochage des jeunes.
Suspendu à une paroi, tête en bas, enveloppé dans son patagium comme dans un manteau, le Grand Rhinolophe a un aspect caractéristique de cocon.
On n’observe aucun dimorphisme sexuel. C’est une espèce nocturne qui vit en petits groupes l’été et hiberne de novembre à avril dans une grotte humide.
La longévité est de 30 ans. Le Grand rhinolophe vit donc très longtemps par rapport à la souris commune d’une taille semblable qui ne vit elle, que deux ans.
Sous-espèces
Il existe six sous-espèces (Thomas[8] 1997, Corbet[9], 1978) :
- R. f. ferrumequinum, en Europe, et Afrique du Nord-Ouest
- R. f. creticum, en Crète
- R. f. irani, Iraq, Iran, Turkménie
- R. f. proximus, Afghanistan, Ouzbékistan Est, Cachemire
- R. f. tragatus Hodgson, 1835 (avant-bras :58-64 mm, tibia > 25 mm), Yunnan, Guizhou, Inde du Nord
- R. f. korai, Corée
- R. f. nippon Temminck, 1835 (avant-bras : 49-59 mm, tibia : 25 mm), vaste bande partant du nord-est de la Chine (Jilin, latitude N 45°) s’étendant vers le sud jusqu’au Sichuan et Yunnan (tropique du Cancer) et l’est jusqu’au Japon.
Reproduction
La maturité sexuelle des femelles se situe entre 2 et 3 ans, celle des mâles, au plus tôt à la fin de la deuxième année[6].
Les accouplements se font principalement en fin d’été et à l’automne, après une période estivale de ségrégation sexuelle totale. À cette époque, les mâles occupent des gîtes d’accouplement déterminés dans des greniers, des grottes ou des galeries, où les femelles viendront les retrouver. Il peut aussi y avoir des accouplements en hiver et au printemps[10]. Les femelles visitent des mâles pour lesquels elles peuvent montrer une certaine fidélité à long terme[11]. La reproduction est essentiellement polygame mais quelques femelles se reproduisent avec le même mâle d’une année sur l’autre et des observations suggèrent que les accouplements seraient davantage polygynes. L’accouplement se fait soit entre individus mâles et femelles issus du même gîte estival soit entre individus issus de gîtes estivaux distincts.
La fécondation est stoppée par la formation d'un bouchon vaginal[12] car les individus rejoignent leur site d'hivernage afin d'hiberner jusque mi-avril. Lorsque les femelles sortent d'hibernation, la fécondation différée se déclenche et la gestation dure de 6 à 8 semaines (selon Arthur et Lemaire[3]) ou plus 10-11 semaines[1]. Les femelles se réunissent en colonie de parturition. Ces colonies qui sont composées d'individus apparentés (grand-mères, mères, filles, cousines...) sont de taille très variable, pouvant aller de 20 individus à près d’un millier. Elles sont parfois associées au Rhinolophe euryale ou au Vespertilion à oreilles échancrées (Myotis emarginatus).
Les mâles sont quant à eux solitaires et utilisent un vaste panel d'habitats.
En France, de mi-juin à fin juillet - début août, les femelles donnent naissance à un seul petit qui n’ouvrira les yeux qu’au 7e jour. La femelle seule s’occupe du petit. Dès le 28e-30e jour, les jeunes apprennent à chasser seuls près du gîte. Mais leurs capacités de vol et d’écholocation sont limitées. Ils sont sevrés vers le 45e jour. Le squelette se développe jusqu’au 60e jour[6].
Les mères accordent une forte importance à l'élevage de leur seul et unique petit annuel et peuvent revenir plusieurs fois la nuit entre des périodes de chasse afin d'allaiter leur progéniture. Des associations ont été constatées avec le Murin à oreilles échancrées (Myotis emarginatus) où des adultes peuvent se placer au milieu des juvéniles de Grand rhinolophe.
Ces observations ont été faites en Europe. En Chine, la répartition va du nord-est (Jilin, Liaoning) avec des hivers beaucoup plus froids que sous les mêmes latitudes en Europe jusqu’au Yunnan traversé par le tropique du Cancer. Dans le sud de la Chine, le Grand rhinolophe est actif toute l’année[5].
Régime alimentaire
Le régime alimentaire varie en fonction des saisons et des pays.
Selon les régions, le régime alimentaire se compose de lépidoptères (30 à 40 %), de coléoptères (25 à 40 %), d’hyménoptères (parasites Ichneumonoidea) (5 à 20 %, les diptères (10 à 20 %), des trichoptères (5 à 10 %)[6] et des orthoptères (sauterelle, criquet)[6].
En Suisse, l’essentiel de la biomasse capturée est constituée de papillons d’avril à septembre, puis de trichoptères de la mi-septembre au début d’octobre. Les coléoptères sont surtout capturés en juillet, les tipules en septembre, les hyménoptères régulièrement en toute saison.
Écholocation
Le Grand rhinolophe émet les cris d’écholocation par ses deux narines. Ils se font sur une fréquence fixe, située entre 79 et 84 kHz. Comme les narines sont entourées d’une feuille nasale en forme de fer à cheval faisant office de pavillon, l’émission sonore est bien mieux focalisée que pour les Vespertilionidés. En produisant des faisceaux étroits dans diverses directions, il peut sonder son environnement grâce à la mobilité de ses oreilles. Toutefois pour pouvoir saisir la modification d’un écho par un mouvement d’oreille, son cri doit durer longtemps et être répété le plus rapidement possible. Mais ces contraintes supposent qu’il puisse entendre les échos de son environnement tout en émettant[7].
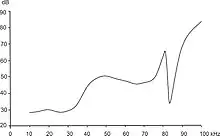
À cette fin, le Grand rhinolophe a développé des adaptations neurologiques remarquables qui lui permettent de percevoir beaucoup mieux certaines fréquences. Sur son audiogramme, on remarque un pic très bas, vers 82 kHz, entouré de zones au contraire de faible sensibilité[13]. À cette fréquence l’oreille est extrêmement sensible alors qu’aux fréquences proches, elle sera très peu sensible. Or quand le rhinolophe est en vol, l’effet Doppler fait que les échos reviennent avec une fréquence plus élevée que celle à laquelle les cris ont été émis. C’est-à-dire qu’en émettant à une fréquence de 77-81 kHz, dans une zone où il est dur d’oreille, il recevra un écho qui grâce à l’effet Doppler se trouvera dans la zone de grande sensibilité. Son oreille ne sera donc pas saturée par le son émis et pourra percevoir clairement l’écho. Dans certaines zones géographiques où seule une sous-espèce est présente, ce pic d'écholocation spécifique est mis à profit pour des études et comptages non invasifs du Grand rhinolophe (exemple de travaux en Tunisie mesurant le pic entre 85 et 88 kHz)[14].
Aire de distribution
L'aire de répartition[3] du Grand rhinolophe s'étend jusqu'à 53°N en Grande-Bretagne, au sud des Pays-Bas et ensuite couvre toute l’Europe méditerranéenne et le nord du Maghreb et vers l'est la Turquie, Iran, Caucase, Turkménie, Ouzbékistan, Afghanistan, Himalaya et jusqu'à la Chine, la Corée et le Japon[4].
Le Grand rhinolophe est présent dans toutes les régions de France, Corse comprise et dans les pays limitrophes[15]
En Europe et en Afrique du Nord, il atteint ses plus hautes densités dans le bassin méditerranéen[16] et ne pénètre guère en Europe centrale. Il atteint ses plus hautes latitudes au Pays de Galles et en Cornouailles, régions qui jouissent d’un climat océanique doux.
À la suite de l’effondrement de ses populations en Europe centrale, sa limite de répartition a régressé vers le sud. En Allemagne, l’espèce a disparu, à l’exception d’une très petite maternité isolée, dans le Haut-Palatinat, et quelques individus en hibernation ici et là, qui passent l’été en Suisse et au Luxembourg.
Populations et habitats
- Effectifs de populations : Les populations de Grand rhinolophe ont subi un fort déclin au cours du XXe siècle. En Grande-Bretagne, les populations comptabilisent environ 5000 individus, en Belgique les populations sont d'environ 200 individus contre moins de 300 individus au Luxembourg. En France, la population a été estimée à 40 000 individus avec des noyaux en Bretagne (8,1 % des effectifs hivernants et 17,4 % des effectifs reproducteurs de l’hexagone[17], soit 37 colonies de reproduction en 2008[18]), dans la Loire et dans le sud de la France. Les populations européennes actuellement les plus importantes sont dans les régions méditerranéennes, les Balkans et la Turquie.
- Habitat hivernal : Pour hiberner, le Grand rhinolophe fréquente des grottes, tunnels, viaducs, anciennes carrières de pierre et des mines de toutes dimensions avec 95 % d'hygrométrie. Il préfère toutefois les vastes gîtes, avec une obscurité totale, où les températures sont comprises entre 5 et 12 °C[6]. Les deux sexes sont accrochés souvent en grappes au plafond.
- Habitat estival : Les femelles de Grand rhinolophe qui se regroupent en colonies de mise-bas, s’installent dans les milieux bâtis chauds (greniers, bâtiments agricoles, combles d'église ou de château) mais aussi caves, accessibles en vol et qui constituent un abri contre les précipitations et les températures froides.
- Gîtes d'étape : Lors des périodes de chasse, le Grand rhinolophe utilise de nombreux gîtes de repos nocturne, ou gîte d'étape, comme les porches, cheminées, viaducs, branches d'arbre, etc.
- Terrains de chasse : Le Grand rhinolophe affecte les milieux bocagers ou les milieux mixtes pour chasser, les haies jouent un rôle important dans ses déplacements et son alimentation car elles recèlent de nombreuses proies et offrent des perchoirs de chasse idéaux. En fin d’été, les pâturages permanents sont recherchés. La proximité de zones aquatiques lui est également favorable.
Le Grand rhinolophe est très fidèle aux gîtes de reproduction et d’hivernage, en particulier les femelles, les mâles ayant un comportement plus erratique.
Il craint les grands rapaces diurnes ou nocturnes. Il est l’objet de la prédation du faucon crécerelle (Falco tinnunculus), de l’épervier d’Europe (Accipiter nisus), de l’effraie des clochers (Tyto alba), de la chouette hulotte (Strix aluco) et du hibou moyen-duc (Asio otus). La présence de chat domestique (Felis catus), de fouine (Martes foina) ou d’effraie des clochers dans un grenier peut être particulièrement néfaste pour les colonies de mise-bas[6].
Activité

Le Grand rhinolophe a bien été étudié en Europe occidentale. Dans cette région, il entre en hibernation de septembre-octobre à avril, suivant les conditions climatiques locales. Dans les régions tropicales, comme au sud de la Chine, il est actif toute l’année.
Le Grand rhinolophe est une espèce strictement nocturne, qui ne chasse pas avant la nuit tombée. Il sort peu pour chasser si le temps est froid, venteux ou pluvieux.
Il est relativement sédentaire : en général, les gîtes d’été et d’hiver sont séparés de 20 à 30 km[6]. Il peut occasionnellement faire un trajet de quelques centaines de kilomètres. En Bulgarie, pour aller chasser en Roumanie, des individus traversent le Danube sur la frontière, qui, à cet endroit mesure jusqu’à 2,5 km de largeur, et reviennent allaiter au milieu de la nuit, pouvant ainsi parcourir plus de 45 km en une nuit[7]. En Tunisie, l'espèce est présente sur l'ensemble du territoire à l'exception de la zone bioclimatique saharienne (même si l'espèce est localisée au Sud jusqu'à Toujane et Tatouine). Elle ne semble pas avoir de préférence concernant l'altitude ou la proximité de la mer : Outres de nombreuses localisations de quelques dizaines d'individus, trois colonies de reproduction ont été identifiées (l'une en bord de mer dans les carrières Fajtun appelées localement "les grottes" au NO d'el Haouariya, et une autre estimée à plus de 300 individus en zone boisée à 1500 m d'altitude, dans le parc naturel el Feija)[14].
Dans les gîtes, il s’accroche au plafond, isolément, en alignement ou en groupe plus ou moins denses. À la tombée de la nuit, le Grand rhinolophe sort de son gîte et vole directement vers des zones de chasse, en suivant préférentiellement les corridors boisés. Il évite les zones ouvertes. Il s’éloigne rarement de plus 2 à 3 km. En Bulgarie, les individus restent dans un rayon de 10 km autour du gîte. Il alterne les phases de chasse et de repos.
Le vol est lent, papillonnant, avec de brèves glissades, généralement à faible hauteur (de 0,3 à 6 m).
Le Grand rhinolophe repère obstacles et insectes par écholocation. La chasse en vol est pratiquée au crépuscule, à un moment où la densité des proies est la plus élevée. En cours de nuit, la chasse à l’affût depuis une branche, devient plus fréquente dans la seconde partie de la nuit[6]. Le Grand rhinolophe émet les ultra-sons par le nez et non par la bouche comme les autres microchiroptères. Son émission sonore est de type fréquence constante avec l'utilisation de l'effet Doppler et des signaux compris entre 81 et 84 kHz, par conséquent sa distance d'émission ultrasonore ne dépasse pas une dizaine de mètres[3].
Statut de protection
- Monde : LR, faible risque. R. ferrumequinum a une grande aire de répartition. Bien qu'il y ait eu des déclins marqués et bien documentés dans certaines régions, l'espèce demeure répandue, abondante et apparemment stable dans d'autres régions.
- Europe du nord-ouest : l’espèce a marqué un déclin prononcé depuis 100 ans dans l’Europe du Nord-Ouest : Royaume Uni, Pays bas, Belgique, Allemagne, Suisse. Le changement d'habitat a probablement été l'une des principales causes de déclin, la conversion des zones boisées et des petits champs en terres agricoles de grande surface étant particulièrement dommageable[15].
- France : quasi menacé. Un recensement partiel en 1997 a compté 25 760 individus répartis dans 1 230 gîtes d’hibernation et environ 8 000 individus dans 196 gîtes estivaux. En Alsace, l’espèce a atteint le seuil d’extinction. La situation est plus favorable dans le Centre, les Ardennes, en Lorraine, Franche-Comté et Bourgogne. Même si l’Ouest de la France, c’est-à-dire la Bretagne, les Pays-de-Loire et le Poitou-Charentes, regroupe encore 50 % des effectifs hivernaux et 30 % des effectifs estivaux, un déclin semble perceptible[6].
Convention de Berne du :
- Annexe 2 : Sont notamment interdits : a) toute forme de capture intentionnelle, de détention et de mise à mort intentionnelle; b) la détérioration ou la destruction intentionnelles des sites de reproduction ou des aires de repos; c) la perturbation intentionnelle de la faune sauvage, notamment durant la période de reproduction, de dépendance et d'hibernation, pour autant que la perturbation ait un effet significatif eu égard aux objectifs de la présente Convention; d) la destruction ou le ramassage intentionnels des œufs dans nature ou leur détention, même vides; e) la détention et le commerce interne de ces animaux, vivants ou morts, y compris des animaux naturalisés, et de toute partie ou de tout produit, facilement identifiables, obtenus à partir de l'animal, lorsque cette mesure contribue à l'efficacité des dispositions de cet article.
Directives européennes du et du concernant la conservation des habitats naturels ainsi que de la faune et de la flore sauvages :
- CE/92/43 - Annexe 2 : Directive Faune-Flore-Habitat, annexe 2 : espèces animales et végétales d'intérêt communautaire dont la conservation nécessite la désignation de zones spéciales de conservation modifiée par la Directive 97/62/CE (espèce dont l'habitat doit être protégé).
- CE/92/43 - Annexe 4 : Directive Faune-Flore-Habitat, annexe 4 : espèce strictement protégée, la capture et la mise à mort intentionnelle est interdite tout comme la perturbation des phases critiques du cycle vital et la destruction de leurs aires de repos et de leurs sites de reproduction.
Convention de Bonn du :
- Accord Chauve-souris : Convention de Bonn sur les espèces migratrices : Accord relatif à la conservation des chauves-souris en Europe : Extraits de l'article 3 sur les obligations fondamentales : 1. Chaque Partie interdit la capture, la détention ou la mise à mort intentionnelle des chauves-souris, sauf lorsqu'il est délivré un permis par son autorité compétente. 2. Chaque Partie identifie, sur le territoire relevant de sa juridiction, les sites qui sont importants pour l'état de la conservation des chauves-souris, notamment pour leur abri et leur protection. En tenant compte au besoin des considérations économiques et sociales, elle protège de tels sites de toute dégradation ou perturbation. Par ailleurs, chaque Partie s'efforce d'identifier et de protéger de toute dégradation ou perturbation les aires d'alimentation importantes pour les chauves-souris.
Menaces sur l'espèce
L'Union international pour la conservation de la nature a classé le Grand Rhinolophe en préoccupation mineure. En Wallonie, l'espèce est jugée vulnérable[19].
Comme toutes les chauves-souris, cette espèce est adaptée à un environnement nocturne. Elle souffre d'une pollution lumineuse croissante, de la fragmentation des habitats, de la diminution de gîtes de mise-bas, du traitement des charpentes et de l'emploi de produits phytosanitaires et vermifuges et de la diminution des populations d’insectes.
Une étude a montré l'impact de l'éclairage direct des individus de cette espèce[20]. L'étude portait sur des colonies de Rhinolophus ferrumequinum, Myotis emarginatus et Myotis oxygnathus vivant dans des bâtiments plus ou moins illuminés ou non éclairés, mais proches les uns des autres. Les chercheurs ont étudié et comparé les dates des naissances, la masse corporelle et la longueur de l'avant-bras de ces chauves-souris et ont constaté que l'éclairage artificiel retardait le développement des jeunes de ces espèces et qu'il pouvait parfois même anéantir toute une colonie. Les petits étaient significativement plus faibles dans les bâtiments illuminés. Les différences de longueur de l'avant-bras et de masse corporelle suggèrent qu'après l'accouchement, le taux de croissance des jeunes est plus faible pour les chauves-souris vivant dans les bâtiments illuminés.
Réservoir naturel de virus
Les chauves-souris fer à cheval sont des réservoirs importants de coronavirus[21]. Depuis les années 2000, plusieurs épidémies de maladies virales sont apparues en Asie où les coronavirus sont passés des chauves-souris à l’homme. À la suite du passage du virus de l’animal à l’homme (ou zoonose), l’épidémie peut se propager à des centaines de milliers de personnes si une transmission interhumaine s’établit et qu'aucun vaccin ne soit disponible.
Dans la base de données Virus-Hôtes DB[22] qui organise les données sur la relation entre les virus et leurs hôtes, on trouve que R. ferrumequinum est l’hôte de 13 types circovirus, cyclovirus, et des coronavirus (Bat CoV 273/2005 [TAX:389166], Bat SARS CoV Rf1/2004 [TAX:347537], BtRf-AlphaCoV/HuB2013 [TAX:1503292], BtRf-AlphaCoV/YN2012 [TAX:1503293], etc.). Dans un article de Shi Zhengli et al. de 2015[23], indiquent que deux nouvelles souches de SL-CoV sont rapportées chez R. ferrumequinum au Yunnan. Il a été suggéré que l’ancêtre du SARS-CoV de la civette (à l'origine de l'épidémie du SRAS ayant débuté près de Canton en Chine) aurait acquis le gène ORF8 de R. ferrumequinum par recombinaison.
Étant donné que les chauves-souris sont connues pour héberger plus de coronavirus que n’importe quelle autre espèce, il est probable que les SARS-CoV, MERS-Cov et SARS-Cov2 ne seront pas les seuls coronavirus à franchir les barrières d’espèces pour causer des épidémies infectieuses.
Recherche
Les chiroptérologues continuent à inventorier les habitats propices à cette espèce, dont avec de nouveaux moyens tels que l'imagerie infrarouge[24]
 Dispositif de vidéosurveillance de la nurserie de Grands Rhinolophes à la citadelle de Montreuil-sur-Mer
Dispositif de vidéosurveillance de la nurserie de Grands Rhinolophes à la citadelle de Montreuil-sur-Mer Caméra infrarouge des Grands Rhinolophes à la citadelle de Montreuil-sur-Mer
Caméra infrarouge des Grands Rhinolophes à la citadelle de Montreuil-sur-Mer
Photos
 Grand Rhinolophe et son petit filmés par une caméra infrarouge à la citadelle de Montreuil-sur-Mer, Pas-de-Calais, France.
Grand Rhinolophe et son petit filmés par une caméra infrarouge à la citadelle de Montreuil-sur-Mer, Pas-de-Calais, France. Grand rhinolophe.
Grand rhinolophe.
Notes
- voir des explications à l’entrée Rhinolophe#Morphologie
Références
- Meyer C., ed. sc., 2009, Dictionnaire des Sciences Animales. consulter en ligne. Montpellier, France, Cirad. consulté en mai 2010.
- (en) Derwent, Thesaurus of agricultural organisms: pests, weeds and diseases, Volume 1. Derwent Publications, Ltd. Éditions CRC Press, 1990. 1529 pages. (ISBN 0-412-37290-8), 9780412372902. Rechercher dans le document numérisé. Consulté en mai 2010.
- Laurent Arthur et Michèle Lemaire, Les Chauves-souris de France Belgique Luxembourg et Suisse, Biotope, coll. « Parthénope », , 576 p. (ISBN 978-2-36662-099-3, présentation en ligne).
- G. Csorba, P. Ujhelyi, N. Thomas, Horseshoe Bats of the World (Chiroptera : Rhinolophidae), Alana Books,
- Andrew T. Smith, Yan Xie, Robert S. Hoffmann, Darrin Lunde, John MacKinnon, Don E. Wilson, W. Chris Wozencraft, A Guide to the Mammals of China, Princeton University Press, , 576 p. (lire en ligne)
- Muséum national d’histoire naturelle, INPN, « Rhinolophus ferrumequinum (Scgreber, 1774), Grand rhinolophe » (consulté le )
- Christian Dietz, Otto von Helversen, Dietmar Nill (traduction Dubourg-Savage), L’encyclopédie des chauves-souris d’Europe et d’Afrique du Nord, delachaux et niestlé, (2007) 2009, 400 p.
- Thomas NM, A systematic review of selected Afro-Asiatic Rhinolophidae (Mammalia : Chiroptera) : an evaluation of taxonomic methodologies, PhD Thesis, Harrison Zoological Museum, Sevenoaks,
- Corbet G.B., The Mammals of Palaearctic Region, Cornell University Press, , 314 p.
- RANSOME RD., (1991) - Greater horseshoe bat. In The handbook of British mammals (ed. G. B. Corbet & S. Harris), p. 88-94. Oxford, UK: Blackwell
- ROSSITER SJ., JONES G., RANSOME RD., FAULKES CG., LE COMBER SC., JONES G., (2005) - Mate fidelity and intra-lineage polygny in greater horseshoe bats — Nat 437, 408-411
- MATTHEWS LH., (1937) - The female sexual cycle of the British horseshoe bats, Rhinolophus ferrumequinum insulanus Barrett- Hamilton and R. hipposideros minutus Montagu—Trans. Zool. Soc. Lond. 23, 224-267
- Neuweiler G, « Neurophysiologische Untersuchungen zum Echo-ortungssystem der Grossen Hufeisennase Rhinolophus ferrumequinum », Z. vergl. Physiol., vol. 67,
- (en) Sebastien J. Puechmaille, Wassim M. Hizem, Benjamin Allegrini, Awatef Abiadh, « Bat fauna of Tunisia: Review of records and new records, morphometrics and echolocation data », Vespertilio Volume 16, , p. 211–239 (lire en ligne)
- Piraccini R., « Greater Horseshoe Bat, Rhinolophus ferrumequinum (UICN) » (consulté le )
- J. Hausser (coord.), Säugetiere der Schweiz / Mammifères de la Suisse : Mammiferi della Svizzera, Springer-Verlag,
- Effectifs et état de conservation des Chiroptères de l’Annexe II de la Directive Habitats-Faune-Flore en France métropolitaine : bilan 2004, rapport Groupes Chiroptères de la Société française pour l'étude et la protection des mammifères, Paris, 31 p.
- Chiffres Contrat Nature Chauve-Souris de Bretagne 2008-2011, Rapport annuel no 4, décembre 2011, 78 p.
- « Liste rouge | Chauves-souris | Espèces | La biodiversité en Wallonie », sur biodiversite.wallonie.be (consulté le )
- sources : 1 et 2(consultées le 01 04 2008)
- Peng Zhou,... Shi Zhengli, « A pneumonia outbreak associated with a new coronavirus of probable bat origin », Nature, vol. 579, (lire en ligne)
- Virus-Host DB (Laboratory of Chemical Life Science, Kyoto University Bioinfomatics Center, Institute for Chemical Research), « Rhinolophus ferrumequinum » (consulté le )
- Ben Hu, Linfa Wang, Zhengli Shi, « Bat origin of human coronaviruses », Virology Journal, vol. 12, no 221, (lire en ligne)
- Parc naturel de Camargue (2014) Techniques d'imagerie au service de la conservation, Conservation et gestion intégrée de 2 espèces de chauve-souris : le grand rhinolophe et le Murin à oreilles échancrées en région méditerranéenne française ; Programme Life+ Chiro Med 2010-2014
Annexes
Articles connexes
Liens externes
- (en) Référence Animal Diversity Web : Rhinolophus ferrumequinum (consulté le )
- (en) Référence Brainmuseum : Rhinolophus ferrumequinum (consulté le )
- (en) Référence Catalogue of Life : Rhinolophus ferrumequinum (Schreber, 1774) (consulté le )
- (fr+en) Référence CITES : espèce Rhinolophus ferrumequinum (Schreber, 1774) (+ répartition) (sur le site de l’UNEP-WCMC) (consulté le )
- (fr) Référence CITES : taxon Rhinolophus ferrumequinum (sur le site du ministère français de l'Écologie) (consulté le )
- (en) Référence Fauna Europaea : Rhinolophus ferrumequinum (Schreber, 1774) (consulté le )
- (fr) Référence INPN : Rhinolophus ferrumequinum (Schreber, 1774) (TAXREF) (consulté le )
- (fr+en) Référence ITIS : Rhinolophus ferrumequinum (Schreber, 1774) (consulté le )
- (en) Référence Mammal Species of the World (3e éd., 2005) : Rhinolophus ferrumequinum Schreber, 1774 (consulté le )
- (en) Référence NCBI : Rhinolophus ferrumequinum (taxons inclus) (consulté le )
- (en) Référence Paleobiology Database : Rhinolophus ferrumequinum Schreber 1774 (consulté le )
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023
- (en) Référence UICN : espèce Rhinolophus ferrumequinum (Schreber, 1774) (consulté le )
- Conseil Chauve-souris blessée
- Contacts chiroptères régionaux en France
- Contrat Nature Grand rhinolophe en Bretagne