Nucléoprotéine
Les nucléoprotéines sont des protéines conjuguées à des acides nucléiques (soit ADN soit ARN)[1]. Les nucléoprotéines typiques comprennent les ribosomes, les nucléosomes et les protéines de la nucléocapside virale.
Structures
Les nucléoprotéines ont tendance à être chargées positivement, facilitant l'interaction avec les chaînes d'acide nucléique chargées négativement. Les structures tertiaires et les fonctions biologiques de nombreuses nucléoprotéines sont comprises[2] - [3]. Les techniques importantes pour déterminer les structures des nucléoprotéines comprennent la diffraction des rayons X, la résonance magnétique nucléaire et la cryomicroscopie électronique.
Virus
Les génomes viraux (ADN ou ARN) sont extrêmement serrés dans la capside virale[4] - [5]. De nombreux virus ne sont donc guère plus qu'une collection organisée de nucléoprotéines avec leurs sites de liaison pointant vers l'intérieur. Les nucléoprotéines virales structurellement caractérisées comprennent la grippe[6], la rage[7], Ebola, Bunyamwera[8], Schmallenberg[8], Hazara[9], la fièvre hémorragique de Crimée-Congo[10], et Lassa[11].
Désoxyribonucléoprotéines
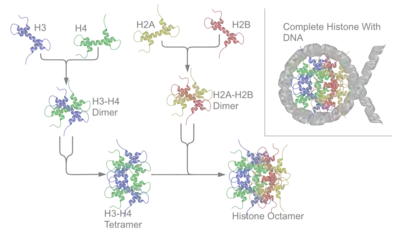
Une désoxyribonucléoprotéine (DNP) est un complexe d'ADN et de protéine[12]. Les exemples prototypiques sont les nucléosomes, des complexes dans lesquels l'ADN génomique est enroulé autour d'amas de huit protéines histones dans les noyaux des cellules eucaryotes pour former la chromatine. Les protamines remplacent les histones lors de la spermatogenèse.
Les fonctions
Les désoxyribonucléoprotéines les plus répandues sont les nucléosomes, dont le composant est l'ADN nucléaire. Les protéines associées à l'ADN sont les histones et les protamines ; les nucléoprotéines résultantes sont situées dans les chromosomes. Ainsi, le chromosome entier, c'est-à-dire la chromatine chez les eucaryotes, est constitué de telles nucléoprotéines[13] - [14].
Dans les cellules eucaryotes, l'ADN est associé à une masse à peu près égale de protéines histones dans un complexe nucléoprotéique très condensé appelé chromatine[15]. Les désoxyribonucléoprotéines de ce type de complexe interagissent pour générer un complexe régulateur multiprotéique dans lequel l'ADN intervenant est bouclé ou enroulé. Les désoxyribonucléoprotéines participent à la régulation de la réplication et de la transcription de l'ADN[16].
Les désoxyribonucléoprotéines sont également impliquées dans la recombinaison homologue, un processus de réparation de l'ADN qui semble être presque universel. Une étape intermédiaire centrale dans ce processus est l'interaction de plusieurs copies d'une protéine recombinase avec de l'ADN simple brin pour former un filament DNP. Les recombinases utilisées dans ce processus sont produites par des archées (recombinase RadA)[17], par des bactéries (recombinase RecA)[18] et par des eucaryotes de la levure à l'homme (recombinases Rad51 et Dmc1)[19].
Ribonucléoprotéines
Une ribonucléoprotéine (RNP) est un complexe d'acide ribonucléique et de protéine de liaison à l'ARN. Ces complexes jouent un rôle essentiel dans un certain nombre de fonctions biologiques importantes, notamment la transcription, la traduction et la régulation de l'expression des gènes [21] et la régulation du métabolisme de l'ARN[22]. Quelques exemples de RNP comprennent le ribosome, l'enzyme télomérase, les ribonucléoprotéines voûtées, la RNase P, le hnRNP et les petits RNP nucléaires (snRNP), qui ont été impliqués dans l'épissage du pré-ARNm (spliceosome) et sont parmi les principaux composants du nucléole[23]. Certains virus sont de simples ribonucléoprotéines, contenant une seule molécule d'ARN et un certain nombre de molécules de protéines identiques. D'autres sont des complexes ribonucléoprotéiques ou désoxyribonucléoprotéiques contenant un certain nombre de protéines différentes et exceptionnellement plus de molécules d'acide nucléique. Actuellement, plus de 2000 RNP sont répertoriées dans la banque de données sur les protéines (PDB) du RCSB[24]. De plus, la base de données d'interface protéine-ARN (PRIDB) possède une collection d'informations sur les interfaces ARN-protéine basée sur des données tirées de la PDB[25]. Certaines caractéristiques communes des interfaces protéine-ARN ont été déduites sur la base de structures connues. Par exemple, les RNP dans les snRNP ont un motif de liaison à l'ARN dans leur protéine de liaison à l'ARN. Les résidus d'acides aminés aromatiques dans ce motif entraînent des interactions d'empilement avec l'ARN. Les résidus de lysine dans la partie hélicoïdale des protéines de liaison à l'ARN aident à stabiliser les interactions avec les acides nucléiques. Cette liaison d'acide nucléique est renforcée par l'attraction électrostatique entre les chaînes latérales positives de lysine et les squelettes négatifs de phosphate d'acide nucléique. De plus, il est possible de modéliser les RNP par calcul[26]. Bien que les méthodes informatiques de déduction des structures RNP soient moins précises que les méthodes expérimentales, elles fournissent un modèle approximatif de la structure qui permet de prédire l'identité d'acides aminés et de résidus nucléotidiques significatifs. Ces informations aident à comprendre la fonction globale de la RNP.
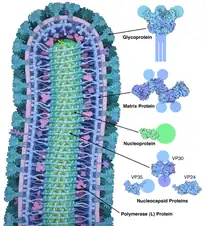
RNP peut également faire référence à des particules de ribonucléoprotéines. Les particules de ribonucléoprotéines sont des foyers intracellulaires distincts pour la régulation post-transcriptionnelle. Ces particules jouent un rôle important dans la réplication du virus de la grippe A[28]. Le génome viral de la grippe est composé de huit particules ribonucléoprotéiques formées par un complexe d'ARN de sens négatif lié à une nucléoprotéine virale. Chaque RNP porte avec lui un complexe ARN polymérase. Lorsque la nucléoprotéine se lie à l'ARN viral, elle est capable d'exposer les bases nucléotidiques qui permettent à la polymérase virale de transcrire l'ARN. À ce stade, une fois que le virus pénètre dans une cellule hôte, il sera prêt à commencer le processus de réplication.
Anticorps anti-RNP
Les anticorps anti-RNP sont des auto-anticorps associés à la maladie mixte du tissu conjonctif et sont également détectés chez près de 40 % des patients atteints de lupus érythémateux. Deux types d'anticorps anti-RNP sont étroitement liés au syndrome de Sjögren : SS-A (Ro) et SS-B (La). Les auto-anticorps anti-snRNP sont appelés anticorps anti-Smith et sont spécifiques du LED. La présence d'un niveau significatif d'anti-U1-RNP sert également d'indicateur possible de MCTD lorsqu'il est détecté en conjonction avec plusieurs autres facteurs[29].
Les fonctions
Les ribonucléoprotéines jouent un rôle de protection. Les ARNm ne se présentent jamais sous forme de molécules d'ARN libres dans la cellule. Ils s'associent toujours aux ribonucléoprotéines et fonctionnent comme des complexes ribonucléoprotéiques[15].
De la même manière, les génomes des virus à ARN à brin négatif n'existent jamais en tant que molécule d'ARN libre. Les ribonucléoprotéines protègent leurs génomes de la RNase[30]. Les nucléoprotéines sont souvent les principaux antigènes des virus car elles possèdent des déterminants antigéniques spécifiques à la souche et au groupe.
Voir également
Notes et références
- (en) MeSH Nucleoproteins
- >Graeme K. Hunter G. K. (2000): Vital Forces. The discovery of the molecular basis of life. Academic Press, London 2000, (ISBN 0-12-361811-8).
- Nelson D. L., Cox M. M. (2013): Lehninger Biochemie. Springer, (ISBN 978-3-540-68637-8).
- Tzlil, Kindt, Gelbart et Ben-Shaul, « Forces and Pressures in DNA Packaging and Release from Viral Capsids », Biophysical Journal, vol. 84, no 3, , p. 1616–1627 (PMID 12609865, PMCID 1302732, DOI 10.1016/s0006-3495(03)74971-6, Bibcode 2003BpJ....84.1616T)
- Purohit, Inamdar, Grayson et Squires, « Forces during Bacteriophage DNA Packaging and Ejection », Biophysical Journal, vol. 88, no 2, , p. 851–866 (PMID 15556983, PMCID 1305160, DOI 10.1529/biophysj.104.047134, Bibcode 2005BpJ....88..851P, arXiv q-bio/0406022)
- (en) Ng, Wang et Shaw, « Structure and sequence analysis of influenza A virus nucleoprotein », Science in China Series C: Life Sciences, vol. 52, no 5, , p. 439–449 (ISSN 1006-9305, PMID 19471866, DOI 10.1007/s11427-009-0064-x, S2CID 610062)
- (en) Albertini, Wernimont, Muziol et Ravelli, « Crystal Structure of the Rabies Virus Nucleoprotein-RNA Complex », Science, vol. 313, no 5785, , p. 360–363 (ISSN 0036-8075, PMID 16778023, DOI 10.1126/science.1125280, Bibcode 2006Sci...313..360A, S2CID 29937744)
- Ariza, Tanner, Walter et Dent, « Nucleocapsid protein structures from orthobunyaviruses reveal insight into ribonucleoprotein architecture and RNA polymerization », Nucleic Acids Research, vol. 41, no 11, , p. 5912–5926 (ISSN 0305-1048, PMID 23595147, PMCID 3675483, DOI 10.1093/nar/gkt268)
- Surtees, Ariza, Punch et Trinh, « The crystal structure of the Hazara virus nucleocapsid protein », BMC Structural Biology, vol. 15, , p. 24 (ISSN 1472-6807, PMID 26715309, PMCID 4696240, DOI 10.1186/s12900-015-0051-3)
- (en) Carter, Surtees, Walter et Ariza, « Structure, Function, and Evolution of the Crimean-Congo Hemorrhagic Fever Virus Nucleocapsid Protein », Journal of Virology, vol. 86, no 20, , p. 10914–10923 (ISSN 0022-538X, PMID 22875964, PMCID 3457148, DOI 10.1128/JVI.01555-12)
- Qi, Lan, Wang et Schelde, « Cap binding and immune evasion revealed by Lassa nucleoprotein structure », Nature, vol. 468, no 7325, , p. 779–783 (PMID 21085117, PMCID 3057469, DOI 10.1038/nature09605, Bibcode 2010Natur.468..779Q)
- (en) MeSH Deoxyribonucleoproteins
- Graeme K. Hunter G. K. (2000): Vital Forces. The discovery of the molecular basis of life. Academic Press, London 2000, (ISBN 0-12-361811-8).
- Nelson D. L., Michael M. Cox M. M. (2013): Lehninger Principles of Biochemistry. W. H. Freeman, (ISBN 978-1-4641-0962-1).
- Harvey Lodish, Molecular Cell Biology
- Echols, « Nucleoprotein structures initiating DNA replication, transcription, and site-specific recombination », The Journal of Biological Chemistry, vol. 265, no 25, , p. 14697–700 (PMID 2203758, DOI 10.1016/S0021-9258(18)77163-9)
- « RadA protein is an archaeal RecA protein homolog that catalyzes DNA strand exchange », Genes Dev., vol. 12, no 9, , p. 1248–53 (PMID 9573041, PMCID 316774, DOI 10.1101/gad.12.9.1248)
- « The importance of repairing stalled replication forks », Nature, vol. 404, no 6773, , p. 37–41 (PMID 10716434, DOI 10.1038/35003501, Bibcode 2000Natur.404...37C, S2CID 4427794)
- « Spontaneous self-segregation of Rad51 and Dmc1 DNA recombinases within mixed recombinase filaments », J. Biol. Chem., vol. 293, no 11, , p. 4191–4200 (PMID 29382724, PMCID 5858004, DOI 10.1074/jbc.RA117.001143)
- Muller, Hutin, Marigold et Li, « A Ribonucleoprotein Complex Protects the Interleukin-6 mRNA from Degradation by Distinct Herpesviral Endonucleases », PLOS Pathogens, vol. 11, no 5, , e1004899 (ISSN 1553-7366, PMID 25965334, PMCID 4428876, DOI 10.1371/journal.ppat.1004899)
- Hogan, Riordan, Gerber et Herschlag, « Diverse RNA-Binding Proteins Interact with Functionally Related Sets of RNAs, Suggesting an Extensive Regulatory System », PLOS Biology, vol. 6, no 10, , e255 (ISSN 1544-9173, PMID 18959479, PMCID 2573929, DOI 10.1371/journal.pbio.0060255)
- Lukong, Chang, Khandjian et Richard, « RNA-binding proteins in human genetic disease », Trends in Genetics, vol. 24, no 8, , p. 416–425 (ISSN 0168-9525, PMID 18597886, DOI 10.1016/j.tig.2008.05.004)
- « Ribonucleoprotein », www.uniprot.org (consulté le )
- « RCSB Protein Data Bank - RCSB PDB », sur web.archive.org, (consulté le )
- Lewis, Walia, Terribilini et Ferguson, « PRIDB: a protein–RNA interface database », Nucleic Acids Research, vol. 39, no Database issue, , D277–D282 (ISSN 0305-1048, PMID 21071426, PMCID 3013700, DOI 10.1093/nar/gkq1108)
- Tuszynska, Matelska, Magnus et Chojnowski, « Computational modeling of protein-RNA complex structures », Methods, vol. 65, no 3, , p. 310–319 (ISSN 1095-9130, PMID 24083976, DOI 10.1016/j.ymeth.2013.09.014, S2CID 37061678)
- Momose, Sekimoto, Ohkura et Jo, « Apical Transport of Influenza A Virus Ribonucleoprotein Requires Rab11-positive Recycling Endosome », PLOS ONE, vol. 6, no 6, , e21123 (ISSN 1932-6203, PMID 21731653, PMCID 3120830, DOI 10.1371/journal.pone.0021123, Bibcode 2011PLoSO...621123M)
- Baudin, Bach, Cusack et Ruigrok, « Structure of influenza virus RNP. I. Influenza virus nucleoprotein melts secondary structure in panhandle RNA and exposes the bases to the solvent. », The EMBO Journal, vol. 13, no 13, , p. 3158–3165 (ISSN 0261-4189, PMID 8039508, PMCID 395207, DOI 10.1002/j.1460-2075.1994.tb06614.x)
- « Mixed Connective Tissue Disease (MCTD) | Cleveland Clinic », my.clevelandclinic.org (consulté le )
- Ruigrok, Crépin et Kolakofsky, « Nucleoproteins and nucleocapsids of negative-strand RNA viruses », Current Opinion in Microbiology, vol. 14, no 4, , p. 504–510 (PMID 21824806, DOI 10.1016/j.mib.2011.07.011)