Cimoliopterus
Cimoliopterus est un genre de ptérosaure qui a vécu au Crétacé supérieur dans ce qui est aujourd'hui l'Angleterre et les États-Unis. Le premier spécimen connu, constitué de la partie avant d'un museau comprenant une partie d'une crête, a été découvert dans le sous-groupe de craie grise du Kent, en Angleterre, et décrit comme la nouvelle espèce Pterodactylus cuvieri en 1851. Le nom spécifique cuvieri rend hommage au paléontologue Georges Cuvier, alors que le genre Pterodactylus était alors utilisé pour de nombreuses espèces de ptérosaures qui ne seraient pas étroitement apparentées aujourd'hui. C'était l'un des premiers ptérosaures à être représentés comme modèles dans le parc de Crystal Palace dans les années 1850. L'espèce a ensuite été attribuée à divers autres genres, notamment Ornithocheirus et Annexeuera. En 2013, l'espèce a été déplacée vers un nouveau genre, sous le nom de Cimoliopterus cuvieri ; le nom générique Cimoliopterus est dérivé des mots grecs pour « craie » et « aile ». D'autres spécimens et espèces ont également été attribués ou synonymes de l'espèce avec divers niveaux de certitude. En 2015, un museau découvert dans la formation de Britton au Texas, aux États-Unis, a été nommé comme nouvelle espèce du genre C. dunni ; le nom spécifique honore son collectionneur, Brent Dunn.
| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Ordre | † Pterosauria |
| Sous-ordre | † Pterodactyloidea |
| Clade | † Ornithocheirae |
| Clade | † Targaryendraconia |
| Famille | † Cimoliopteridae |
Espèces de rang inférieur
- † Cimoliopterus cuvieri Bowerbank, 1851 (espèce type)
- † Cimoliopterus dunni Myers, 2015
On estime que C. cuvieri avait une envergure de 3,5 m et que C. dunni était d'une taille similaire. Cimoliopterus peut être distingué des ptérosaures apparentés par des caractéristiques telles qu'une crête prémaxillaire commençant vers l'arrière du museau, une crête sur le palais qui s'étend vers l'avant jusqu'à la troisième paire d'alvéoles dentaires, et par l'espacement et les proportions des alvéoles dentaires. Contrairement aux ptérosaures similaires, le bout du museau n'est que subtilement élargi sur les côtés. C. cuvieri et C. dunni diffèrent l'un de l'autre par divers détails dans la configuration de ces caractéristiques ; par exemple, la crête de C. cuvieri commence par l'alvéole de la septième dent, alors que celle de C. dunni commence à la quatrième. Les genres apparentés les plus connus étaient des ptérosaures assez grands, avec des crânes proportionnellement grands, de longues mâchoires et des rangées de dents, souvent avec de grandes crêtes arrondies à l'avant des mâchoires. Les dents à l'avant des mâchoires étaient grandes et recourbées ; plus en arrière, les dents étaient plus petites, légèrement recourbées et bien espacées. En tant que ptérosaures, les Cimoliopterus auraient été recouverts de pycnofibres (filaments ressemblant à des cheveux) et auraient des membranes alaires extensibles, qui étaient distendues par de longs doigts alaires.
Bien que longtemps considéré comme un ornithocheirien, les affinités de C. cuvieri n'étaient pas claires en raison de sa nature fragmentaire et d'autres ptérosaures anglais, jusqu'à ce que des parents plus complets soient signalés au Brésil dans les années 1980. Cimoliopterus a été transféré dans la famille des Cimoliopteridae au sein du clade Targaryendraconia en 2019, son plus proche parent étant Camposipterus. Le fait que C. cuvieri et C. dunni, originaires respectivement d'Angleterre et d'Amérique du Nord, étaient si similaires bien qu'ils vivent sur les côtés opposés de l'océan Atlantique Nord, indique qu'ils sont devenus moins isolés les uns des autres que les autres groupes d'animaux parce qu'ils pouvaient voler. Ce type de ptérosaure était probablement adapté pour le vol océanique à longue distance, ce qui est également soutenu par leurs fossiles trouvés principalement dans des environnements marins. Bien qu'ils aient pu maîtriser l'eau, leurs capacités terrestres étaient limitées en raison de leurs membres postérieurs courts mais de leurs membres antérieurs longs. Ils se nourrissaient peut-être en volant en plongeant leurs longues mâchoires dans l'eau et en attrapant des poissons avec leurs dents imbriquées. Alors que certains ont suggéré que les crêtes prémaxillaires étaient utilisées pour stabiliser les mâchoires lorsqu'elles étaient immergées dans l'eau pendant l'alimentation, elles peuvent avoir été utilisées comme structures de parade à la place.
Histoire


L'histoire taxonomique de Cimoliopterus et des ptérosaures apparentés du Crétacé d'Angleterre a été décrite comme déroutante par les chercheurs modernes sur les ptérosaures, car nombre d'entre eux ont été décrits à partir de restes très fragmentaires et nommés à l'aide de schémas nomenclatural alambiqués de l'ère victorienne[1] - [2] - [3]. Avant ces découvertes, on pensait que de nombreux os de ptérosaure creux et à paroi mince trouvés en Angleterre appartenaient à des oiseaux[4]. En 1851, le naturaliste britannique James Scott Bowerbank a décrit un grand museau de ptérosaure qu'il avait obtenu, qui a été trouvé dans le Lower Culand Pit dans ce qu'on appelle maintenant le sous-groupe Grey Chalk à Burham, Kent, dans l'Angleterre du sud-est. Des fossiles de ptérosaures avaient été découverts plus tôt dans la même fosse, y compris la partie antérieure de certaines mâchoires que Bowerbank avait utilisées comme base pour l'espèce Pterodactylus giganteus en 1846, ainsi que d'autres ossements. Sur la base du nouveau museau, Bowerbank a nommé l'espèce Pterodactylus cuvieri ; à cette époque, le genre Pterodactylus (nommé à l'origine en 1815 sur la base d'un spécimen bavarois) était utilisé pour de nombreuses espèces de ptérosaures que l'on croyait maintenant éloignées les unes des autres[5] - [6] - [7]. Le nom spécifique honore le paléontologue français Georges Cuvier, qui avait reconnu que les ptérosaures étaient des reptiles volants[4] - [8].
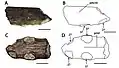
Le museau dont Bowerbank a fait le spécimen holotypique (sur lequel le nom scientifique est basé) de P. cuvieri se compose de l'avant de la mâchoire supérieure, y compris une partie d'une crête prémaxillaire (une crête sur le prémaxillaire, l'os le plus en avant de la mâchoire supérieure), et est catalogué aujourd'hui comme NHMUK PV 39409 au Musée d'histoire naturelle de Londres. Il était à l'origine rapporté qu'il préservait une seule dent dans la première alvéole droite (à l'avant du museau), mais elle avait disparu lorsque l'holotype a été examiné dans les années 2000. Deux dents complètes ont également été signalées à l'origine comme étant conservées dans le même bloc de craie que le museau. Bowerbank pensait également que certains gros os de trois autres collections pouvaient appartenir à la même espèce, à P. giganteus, ou à une troisième espèce possible[5] - [9] - [10]. Le biologiste britannique Richard Owen a attribué un os long à P. cuvieri en 1851 (aujourd'hui catalogué sous le nom de NHMUK PV 41637) en fonction de sa taille et du caractère de sa surface, et l'a identifié comme une phalange possible du doigt allongé de l'aile[10] - [11].

Dans les années 1850, l'artiste britannique Benjamin Waterhouse Hawkins a créé des sculptures grandeur nature d'animaux préhistoriques pour le Parc Crystal Palace à Londres, sous la supervision d'Owen. Parmi eux se trouvaient deux espèces de ptérosaures, dont deux sculptures de P. cuvieri en béton à ossature de fer et deux statues plus petites de Pterodactylus bucklandi[12] - [13] - [14]. Les modèles de Hawkins ont été les premiers à représenter des ptérosaures. Incidemment, il pensait que ces animaux étaient les dragons originaux, comme dans la légende de Saint George[14]. Les plus grandes statues de P. cuvieri sont aujourd'hui en mauvais état en raison de leur nature délicate et du vandalisme, et les plus petites ont été perdues. Le paléontologue britannique Mark Witton a déclaré en 2019 que l'anatomie des sculptures était probablement basée sur le (alors) plus complètement connu P. antiquus. Ils étaient représentés avec des écailles (bien que les ptérosaures soient maintenant connus pour avoir une couverture en forme de cheveux, une caractéristique déjà réalisée par certains scientifiques à l'époque) et des proportions d'oiseaux telles que de petites têtes et de gros torses, comme c'était la coutume à l'époque. . Une sculpture est correctement représentée dans une pose quadrupède, et avec un pied à plat sur le sol[13] - [15].
En 1869, le paléontologue britannique Harry Govier Seeley a placé P. cuvieri dans le nouveau genre Ptenodactylus avec d'autres ptérosaures anglais connus principalement des mâchoires supérieures, dans un index de spécimens du musée Woodwardian. Il a noté que ces noms étaient provisoires, uniquement destinés à la commodité des étudiants utilisant le musée, et pas nécessairement comme noms pour ces espèces[16]. En 1870, Seeley a placé l'espèce dans le genre Ornithocheirus, qu'il avait initialement utilisé pour d'autres espèces dans son index de 1869, et a estimé qu'un os dentaire (os antérieur de la mandibule) qu'il a répertorié appartenait à O. cuvieri[17]. En 1874, Owen a attribué l'espèce au genre Coloborhynchus[18]. Le paléontologue britannique Reginald Walter Hooley a utilisé le nom O. cuvieri dans sa révision de 1914 d'Ornithocheirus, en utilisant la terminologie de Seeley[19]. En 1922, le naturaliste autrichien Gustav von Arthaber a déploré que la littérature scientifique ait accepté les nombreux noms d'Ornithocheirus qui n'avaient été mentionnés que dans le catalogue pour étudiants de Seeley. À son avis, les noms n'étaient d'aucune utilité sans une illustration des spécimens sur lesquels ils étaient basés, ou mieux encore, une reconstruction complète du squelette concerné. Pour cette raison, il a fourni un dessin du crâne d'O. cuvieri (parmi d'autres espèces), l'une des rares espèces pour lesquelles le matériel de mâchoire connu a prouvé sa validité[20]. En 1924, les interprétations de von Arthaber ont été critiquées par le paléontologue hongrois Franz Nopcsa von Felső-Szilvás, qui a trouvé certaines des reconstructions du crâne « sans valeur »[21].
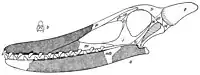

O. cuvieri et de nombreux autres ptérosaures anglais ont été conservés dans le genre Ornithocheirus pendant la majeure partie du 20e siècle[2]. En 1987, le paléontologue allemand Peter Wellnhofer a décrit le nouveau ptérosaure à crête Tropeognathus de la formation de Santana au Brésil et a noté les similitudes entre celui-ci et d'autres taxons brésiliens nouvellement décrits tels que Anhanguera avec les taxons anglais qui étaient basés sur des museaux fragmentaires, tels que les différentes espèces attribué à Ornithocheirus. Il a conclu que si l'apparition des taxons anglais avait longtemps été une énigme (conduisant par exemple aux reconstructions inhabituelles de von Arthaber), la découverte des espèces brésiliennes apparentées et bien mieux conservées a rendu cela plus clair, montrant que les espèces anglaises avaient également des crêtes prémaxillaires au bout de grands et longs crânes[1] - [22]. Quand Anhanguera a été décrit en 1985, on pensait qu'il s'agissait du seul ptérosaure avec une telle crête connue jusqu'à ce point (alors appelée crête sagittale)[23]. En 1991, Wellnhofer a déclaré que le genre Ornithocheirus était devenu une « poubelle » d'espèces de différents pays, attribuée sur la base de caractères insuffisants, et avait besoin d'une définition plus claire, et comprenait peut-être plusieurs genres distincts[24].
Le paléontologue britannique S. Christopher Bennett a déclaré en 1993 que le spécimen holotype d'O. cuvieri était le premier os de la phalange de l'aile droite mentionné par Owen en 1851, et l'a cité pour le nom sans autre explication[25]. En 1995, la paléontologue russe Natalia N. Bakhurina et le paléontologue britannique David M. Unwin ont attribué une symphyse mandibulaire partielle (la partie reliant les deux moitiés de la mâchoire inférieure) de Russie à cf. Anhanguera cuvieri (du latin "confer", appartenant peut-être à cet animal ou à un animal apparenté), utilisant ainsi une Combinatio nova[26]. Unwin et ses collègues ont attribué des éléments squelettiques de ptérosaures de la formation de Toolebuc en Australie à Anhanguera ?cuvieri en 2000[27]. En 2001, Unwin a également utilisé la combinaison A. cuvieri et a cité Bowerbank pour le nom, avec le museau comme holotype. Il a attribué 23 fragments de mâchoire de la formation de Cambridge Greensand à A. cuvieri et l'a considéré comme le ptérosaure le plus commun de cette formation. Unwin a conclu que les espèces O. brachyrhinus, O. colorhinus, O. dentatus, O. denticulatus, O. enchorhynchus, O. scaphorhynchus et O. xyphorhynchus étaient des synonymes juniors d'A. cuvieri. Il a également considéré une espèce nommée à l'origine Pterodactylus fittoni par Owen en 1859 en tant que membre d'Anhanguera[2]. En 2011, le paléontologue brésilien Alexander W. Kellner et ses collègues ont fait du matériel australien attribué à A. ?cuvieri la base d'un nouveau genre, Aussiedraco, et ont déclaré qu'ils ne voyaient aucune raison de placer O. cuvieri à Anhanguera[28].
Nouveau genre et espèce assignée
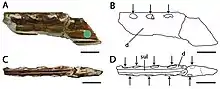
En 2013, les paléontologues brésiliens Rodrigues (d) et Alexander Kellner ont passé en revue les espèces qui avaient jusqu'à présent été placées dans le genre Ornithocheirus (qu'ils ont restreint à son espèce type, O. simus), ainsi que d'autres ptérosaures anglais du Crétacé. Ils ont conclu qu'A. cuvieri différait suffisamment de ses parents pour être placé dans un nouveau genre monotypique ; Cimoliopterus. Ce nom générique combine les mots grecs Kimolia (Κιμωλία), qui signifie « craie », et pteron, (πτερόν) qui signifie « aile ». Rodrigues et Kellner ont également découvert que si les espèces P. fittoni, O. brachyrhinus et O. enchorhynchus avaient diverses caractéristiques en commun avec C. cuvieri et ne pouvaient donc pas être exclues de ce genre, elles étaient trop fragmentaires pour lui être attribuées définitivement et étaient considérés comme des nomina dubia (noms douteux). Ils considéraient que d'autres espèces attribuées à A. cuvieri par Unwin en 2001 étaient douteuses ou appartenant à d'autres taxons. Ils ont également considéré le spécimen russe attribué à cf. A. cuvieri en 1995 sans lien de parenté[29]. En 2019, le paléontologue brésilien Rodrigo V. Pêgas et ses collègues ont suggéré que le spécimen type de Lonchodectes (un genre douteux à l'origine connu sous le nom de Pterodactylus compressirostris) représenté par une symphyse mandibulaire partielle du Chalk Group, pourrait représenter la même espèce que C. cuvieri. Ils ont averti qu'il est impossible de confirmer tant que le crâne et le matériel de la mandibule associés ne sont pas trouvés[30].
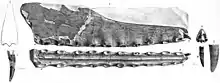
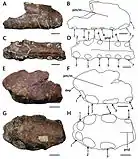
En 2013, le collectionneur amateur américain de fossiles Brent Dunn a découvert un fragment de museau de ptérosaure dans la formation de Britton près du Lewisville Lake, au nord-ouest de Dallas, Texas, États-Unis. Il a fait don du spécimen au Shuler Museum of Paleontology de l'Université méthodiste du Sud, où il est catalogué sous le nom de SMU 76892. En 2015, le paléontologue américain Timothy S. Myers a décrit le spécimen et en a fait l'holotype de la nouvelle espèce Cimoliopterus dunni ; le nom spécifique honore Dunn, décédé en 2013. Le spécimen holotype de C. dunni se compose de la partie antérieure d'un museau, y compris une partie d'une crête prémaxillaire. Le fossile était initialement recouvert d'une fine couche d'argile rouge, qui a été préparée. Le fossile préservé en trois dimensions ne semble pas avoir subi l'écrasement extrême souvent observé dans les fossiles de ptérosaure, bien qu'une partie de la partie postérieure ait été déformée par le compactage. C. dunni n'était que le troisième ptérosaure à dents connu du Crétacé d'Amérique du Nord[7] - [31].
Description

Parce que les fossiles sont incomplets, la taille de Cimoliopterus est difficile à estimer[30]. En extrapolant à partir des restes d'espèces de ptérosaures plus complètes, ainsi que de divers os appartenant peut-être à C. cuvieri, Bowerbank a estimé une envergure d'environ 5,1 mètres en 1851. C'était plus grand que les estimations pour le plus tôt nommé P. giganteus (maintenant dans le genre Lonchodraco), qu'il considérait comme « gigantesque » pour un ptérosaure[5] - [32]. Plus tard la même année, Owen a qualifié C. cuvieri de « vraiment gigantesque », par rapport à P. giganteus, qui, selon lui, était donc nommé de manière inexacte[33]. En 2001, en comparant le spécimen d'holotype de C. cuvieri avec l'Anhanguera plus complet, Unwin a estimé une envergure d'environ 3,5 mètres. Il a estimé qu'un spécimen assigné, CAMSM B54.431, avait une envergure allant jusqu'à 5 mètres[2]. En 2013, ce spécimen - à l'origine une extrémité du museau syntype d'Ornithocheirus colorhinus - a été provisoirement attribué à Camposipterus par Rodrigues et Kellner[9]. En 2015, Myers a déclaré que les holotypes de C. cuvieri et C. dunni appartenaient à des individus de taille similaire[7]. C. dunni aurait été de taille moyenne pour un ptérosaure, avec une envergure d'environ 1,8 mètre, selon un communiqué de presse accompagnant sa description[31]. En 2019, Pêgas et ses collègues se sont abstenus d'estimer l'envergure de ces espèces représentées par du matériel trop fragmentaire[30].
Les genres apparentés les plus connus étaient des ptérosaures assez grands, avec des crânes proportionnellement grands, de longues mâchoires et des rangées de dents, et souvent avec de grandes crêtes arrondies à l'avant des mâchoires. Les dents à l'avant des mâchoires étaient grandes et recourbées, et plus en arrière, les dents étaient plus petites, légèrement recourbées et bien espacées. Une grande partie du squelette aurait été pneumatisée (remplie de sacs aérien qui l'allégeraient). Les cous étaient proportionnellement longs et les torses relativement petits. Les membres antérieurs étaient proportionnellement énormes par rapport aux jambes, avec de très longs doigts ailés[1]. En tant que ptérosaure, Cimoliopterus aurait été recouvert de pycnofibres (filaments ressemblant à des cheveux) et avait de vastes membranes alaires, qui étaient distendues par les doigts de l'aile[34].
Rodrigues et Kellner ont fourni un diagnose unique (une liste de caractéristiques distinguant un taxon de ses proches) pour le genre Cimoliopterus et l'espèce C. cuvieri en 2013, que Myers a modifié en 2015 en incluant C. dunni. Les museaux holotypes des deux espèces de Cimoliopterus partagent des caractéristiques qui les distinguent des autres ptérosaures ptérodactyloïdes (ou à queue courte) comme une crête prémaxillaire qui commence vers l'arrière sur le museau et que la crête palatine (qui longeait le milieu du palais) s'étendait vers l'avant jusqu'à la troisième paire d'alvéoles dentaires. Chez les deux espèces, les deuxième et troisième alvéoles dentaires sont de taille similaire et plus grandes que la quatrième. L'espacement entre les cavités est irrégulier, celles vers l'avant étant plus rapprochées et celles vers l'arrière plus largement séparées. Il y a trois alvéoles par 3 cm de marge de la mâchoire vers l'avant de la mâchoire chez C. dunni, tandis que chez C. cuvieri il y a presque trois alvéoles par 3 cm. Cependant, vers l'arrière de la mâchoire, il y a deux alvéoles tous les 3 cm chez les deux espèces. Le palais est courbée vers le haut[7] - [9]. Les extrémités du museau de C. cuvieri et C. dunni ne s'étendent que subtilement d'un côté à l'autre, contrairement aux expansions en forme de cuillère observées chez de nombreux autres ptérodactyloïdes dentés, tels que Anhanguera, Coloborhynchus et Ornithocheirus. L'expansion latérale subtile sur le bout du museau de Cimoliopterus résulte du fait que la troisième paire de cavités est plus grande que la quatrième paire, semblable à ce qui peut être vu sur la mâchoire inférieure d'Aetodactylus. L'expansion subtile est si faiblement développée qu'elle a parfois été considérée comme absente[7] .
Cimoliopterus cuvieri


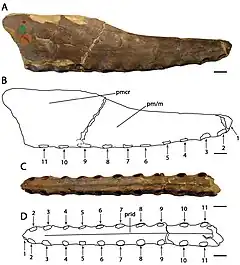
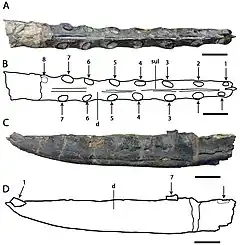
L'holotype de C. cuvieri est composé des 18 cm de devant du museau, représentés principalement par les prémaxillaires, dont la crête prémaxillaire. Onze alvéoles dentaires sont conservées de chaque côté[5] - [9]. La partie conservée du museau est étroite. C. cuvieri se distingue par le fait que la crête prémaxillaire est placée vers l'arrière du museau par la septième paire de cavités, mais avant la fenêtre naso-antéorbitaire (une grande ouverture devant l'œil qui combinait la fenêtre antéorbitaire avec la narine osseuse)[9]. C. cuvieri a un museau bas par rapport à Ornithocheirus et possède également une première paire d'alvéoles dentaires orientée vers l'avant, contrairement à ce genre. De plus, C. cuvieri peut être distingué du genre Anhanguera car il n'a pas d'expansion à l'avant du museau, caractéristique par ailleurs présente dans la famille des Anhangueridae. C. cuvieri n'a pas non plus ses quatrième et cinquième paires d'alvéoles dentaires plus petites que ses troisième et sixième, ce qui est une caractéristique clé d'Anhanguera[7] - [9] - [30].
Bowerbank et Owen ont décrit et figuré les dents maintenant perdues trouvées avec l'holotype de C. cuvieri en 1851. La droite des deux cavités les plus à l'avant contenait une dent nouvellement éclatée (émergée par les gencives), qui dépassait d'environ un tiers de pouce vers le bas et vers l'avant sous un angle oblique. La cinquième alvéole à droite et la huitième à gauche contenaient des dents en herbe qui ne dépassaient pas des alvéoles, se trouvant près des parois internes des alvéoles des dents complètement sorties. Les deux dents déplacées complètement développées étaient légèrement incurvées, quelque peu comprimées, et leur largeur diminuait progressivement depuis les bases ouvertes jusqu'à leurs extrémités. Les pointes ont été rompues, révélant qu'elles étaient composées de dentine compacte et dure, recouverte d'une fine couche d'émail. De fines crêtes longitudinales (ou stries) de longueur inégale, avec de larges intervalles marquent l'émail. La plus grande des dents déplacées mesurait plus de 3 cm, tandis que l'autre mesurait moins de 3 cm et se trouvait près de la troisième paire d'alvéoles dentaires[5] - [7] - [10].
D'autres espèces de ptérosaures ont été considérées comme similaires à C. cuvieri, mais la plupart sont connues de manière trop fragmentaire pour lui être incontestablement attribuées ou synonymes de lui. Par exemple, Pterodactylus fittoni partage avec C. cuvieri une pointe de museau basse, l'absence d'expansion frontale au bout du museau, un palais courbé vers l'arrière, ainsi qu'une crête palatine s'étendant vers l'arrière jusqu'à la troisième paire de les alvéoles dentaires, bien que cette dernière caractéristique ne soit partagée avec C. cuvieri que si la première paire d'alvéoles dentaires conservée chez P. fittoni est sa deuxième paire. La hauteur du museau de P. fittoni peut être différenciée de celle de C. cuvieri, dont la pointe est également plus large que haute ; cette dernière différence est peut-être due à la fracture, cependant, et l'espèce ne peut pas être attribuée de manière indiscutable. Une autre espèce, Ornithocheirus brachyrhinus, partage un certain nombre de caractéristiques avec C. cuvieri, notamment un palais incurvé, l'extrémité antérieure étant plus haute que large, l'absence d'expansion latérale à l'avant du museau et l'absence de crête à l'avant de le museau. La structure de l'extrémité du museau d'O. brachyrhinus ressemble totalement à celle de C. cuvieri, suggérant que O. brachyrhinus est attribuable à cette espèce. Pour autant, l'holotype d'O. brachyrhinus est fragmentaire, et il est donc impossible de conclure s'il possède une crête située vers l'arrière, ou s'il présente la variation de taille des alvéoles dentaires propre à C. cuvieri[2][35].
D'autres espèces qui ont été considérées comme des synonymes de C. cuvieri, comme O. dentatus et O. enchorhynchus, diffèrent, la première ayant des alvéoles dentaires plus petites qui sont rapprochées. Ce dernier, bien qu'assez similaire à C. cuvieri (partageant des caractéristiques telles que l'absence de crête positionnée vers l'arrière, le palais courbé vers l'arrière, la première paire de dents tournée vers l'avant, ainsi que l'absence d'expansion avant), est fragmentaire comme O. brachyrhinus, ce qui rend difficile la comparaison avec les autres ptérosaures. O. scaphorhynchus a également été provisoirement synonyme de C. cuvieri, mais le caractère incomplet de cette espèce rend difficile son attribution claire à un genre. Les marges des alvéoles dentaires chez O. scaphorhynchus sont mal conservées, mais elles sont différentes du modèle d'espacement présent chez C. cuvieri[2][35].
Cimoliopterus dunni

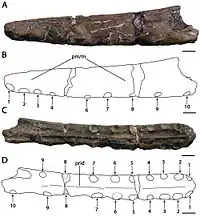
Le museau holotype de C. dunni est composé des 18,5 cm avant du museau, comprenant le prémaxillaire et le maxillaire (y compris une crête prémaxillaire en grande partie intacte), rompus juste derrière la 13e paire d'alvéoles dentaires. L'os cortical (la couche externe dure des os) est bien conservé, bien qu'il existe plusieurs régions où il est endommagé ou manquant. Le nombre total d'alvéoles dentaires conservées dans l'échantillon est de vingt-six. C. dunni se distingue par la crête prémaxillaire commençant à la quatrième paire d'emboîtures, contrairement à celle de C. cuvieri, et la crête est légèrement incurvée vers le haut, formant un bord antérieur concave[7]. La crête est entièrement soudée à la mâchoire, ce qui indique que l'individu n'était pas un jeune[31]. Le bord supérieur de la crête de C. dunni semble descendre modérément juste avant la partie brisée, ce qui suggère que la crête était symétrique d'avant en arrière lorsqu'elle était vue de côté. Si la crête était symétrique, la crête complète aurait eu une longueur d'environ 15 à 16 centimètres. La hauteur maximale de la crête est de 38 millimètres, ce qui est au-dessus de la neuvième et de la dixième paire d'alvéoles dentaires. Alors que les caractéristiques liées à la crête doivent être utilisées avec prudence lors de l'identification des espèces, car elles peuvent être liées au stade de croissance ou au dimorphisme sexuel, la différence de forme de crête entre C. dunni et C. cuvieri n'est probablement pas liée à l'âge, puisque les holotypes représentent individus de même taille[7].
Le bout du museau de C. dunni est émoussé. Il se distingue par le fait que la surface antérieure de la mâchoire est tournée vers l'avant et vers le bas, formant un angle de 45° par rapport à la partie antérieure du palais. En regardant l'extrémité avant du museau de C. dunni sous le palais, il se dilate à mesure qu'il recule jusqu'à une largeur maximale de 1,6 centimètre au-dessus de la troisième paire d'alvéoles dentaires. Il se rétrécit soudainement à une largeur minimale de 1,5 centimètre (0,59 in) au niveau de la quatrième paire de cavités. La largeur du museau continue d'augmenter vers l'arrière jusqu'à atteindre un maximum de 1,8 à 1,9 cm au niveau de la partie cassée du bord postérieur de la crête. Un angle de 8° par rapport au méplat de la partie postérieure du palais est présumé basé sur la réflexion de sa partie antérieure vers l'arrière. Un point d'inflexion (le point où la surface change) proche du niveau des alvéoles de la huitième dent peut également être vu[7] .
C. dunni se distingue de C. cuvieri en ce que la première paire d'alvéoles dentaires est dirigée plus vers le bas que vers l'avant. Les espaces entre les alvéoles mesurent un maximum de 11,5 millimètres vers l'avant et un minimum de 1,6 millimètres vers l'arrière. Comme seule une petite dent de remplacement est conservée chez C. dunni, la texture de ses dents est inconnue. Bien que la couronne (la partie recouverte d'émail) de cette dent soit dépourvue de stries, il est possible qu'elles soient apparues sur des dents plus développées. À moins de 1 millimètre de haut, une étroite crête palatine s'étend vers l'avant à partir de la partie cassée à l'extrémité arrière de la crête prémaxillaire[7] .
Classification
Anciens schémas

L'histoire taxonomique de l'espèce type de Cimoliopterus, C. cuvieri, est compliquée, bien que les ptérosaures anglais maintenant connus pour avoir été de proches parents aient toujours été considérés comme similaires. Lorsqu'il a été décrit pour la première fois, Bowerbank a attribué C. cuvieri (alors placé dans Pterodactylus) à un groupe d'« espèces à long nez », avec l'espèce Pterodactylus longirostris sur la base de proportions de museau similaires[5]. Plus tard, Owen a attribué P. cuvieri au Dentirostres, un groupe caractérisé par des mâchoires contenant des dents à leurs extrémités, un anneau sclérotique (anneau d'os qui entoure les yeux), leur scapula (omoplate) et coracoïde (structure en forme de crochet dans l'épaule ) n'étant pas reliés les uns aux autres, ainsi qu'ayant une courte queue mobile. P. cuvieri a été classé dans ce groupe en raison de l'extrémité de sa mâchoire supérieure, ce qui était suffisant pour démontrer qu'il avait une paire d'alvéoles dentaires proches de l'extrémité[10].
En 1870, Seeley attribua P. cuvieri (sous le nom Ornithocheirus cuvieri dans sa classification) à un groupe nommé Ornithocheirae, qui ne comprenait que le genre Ornithocheirus, dont l'espèce a été diagnostiquée comme ayant des dents prolongées devant le museau et un palais avec une crête longitudinale (dans le sens de la longueur)[17]. En 1914, Hooley a passé en revue le genre Ornithocheirus et sa taxonomie, le divisant en groupes numérotés avec un nom générique (genre) désigné. Hooley a suivi Seeley en classant P. cuvieri comme une espèce d'Ornithocheirus, attribuant également les espèces O. colorhinus et O. nasutus (maintenant considérées comme des espèces de Camposipterus[9]), ainsi que les espèces O. brachyrhinus, O. dentatus, O denticulatus, O. enchorhynchus, O. fittoni et O. oxyrhinus, en plus des espèces O. sedgwicki (maintenant considérée comme l'espèce type d'Aerodraco[36], ou encore une espèce de Camposipterus[9][37]) et également O. polyodon. Hooley a nommé ce groupe « Groupe n° 1 » et lui a donné le nom de genre Ornithocheirus. Il a ensuite attribué ce genre à une sous-famille appelée Ornithocheirinae, qui appartenait à une famille appelée Ornithocheiridae. Toutes les espèces du genre Ornithocheirus ont été diagnostiquées comme ayant des becs comprimés latéralement, avec la pointe plus ou moins obtus (avec un angle supérieur à 90°), leur palais étant légèrement recourbé vers le haut et tourné vers l'arrière (ce qui a fait que leurs dents de devant pointent directement vers l'avant ), et ayant une crête longitudinale en bouche. Leurs dents étaient presque circulaires et les marges de leurs alvéoles s'élevaient au-dessus du palais[19].
Schémas du XXIe siècle

En 2001, Unwin considérait A. cuvieri comme un membre des Ornithocheiridae, avec de nombreuses espèces du Crétacé anglais et brésilien, et suggéra que certaines espèces de ptérosaures de ces pays appartenaient aux mêmes genres, comme Anhanguera, Ornithocheirus et Coloborhynchus[2]. En 2013, Witton a décrit l'état de la classification des ornithocheiridés comme tristement controversé et confus, avec des schémas concurrents utilisés par différents groupes de paléontologues. Un groupe a utilisé le terme Ornithocheiridae, tandis qu'un autre a utilisé Anhagueridae, et il n'y avait pas d'accord sur le nombre de genres et d'espèces[1]. Dans sa description par Rodrigues et Kellner en 2013, Cimoliopterus, qui ne comprenait que C. cuvieri, a été attribué au groupe Pteranodontoidea, bien qu'en position incertaine (incertae sedis), avec lui et « Ornithocheirus » polyodon formant un groupe frère au nouveau clade Anhanguera. Ils n'ont pas pu confirmer que les Ornithocheiridae étaient un groupe monophylétique (naturel) et l'ont limité à Ornithocheirus simus seul[38]. En 2014, dans une analyse phylogénétique (l'étude du développement évolutif d'une espèce ou d'un groupe) menée par le paléontologue chinois Xiaolin Wang et ses collègues, il a été découvert que Cimoliopterus était étroitement lié au ptérosaure Ikrandraco, qui à l'époque, avec les ptérosaures appartenant aux Anhangueria, étaient les seuls connus à avoir eu des crêtes en forme de lame sur leurs mâchoires[39].
Dans sa description de C. dunni, Myers a utilisé une version modifiée de l'ensemble des données d'une analyse phylogénétique plus ancienne qu'il a menée avec le paléontologue Brian Andres en 2013[7] - [40]. La disposition des cladogrammes résultants (« arbres généalogiques » montrant des interrelations) dépendait de la façon dont le la texture incertaine des dents de C. dunni a été interprétée ; s'il était codé comme « ambigu », C. dunni devenait le taxon frère d'un clade composé de C. cuvieri et Aetodactylus halli, et s'il était codé comme strié, C. dunni devenait le taxon frère de C. cuvieri, avec A .halli comme taxon frère de ce clade. À son tour, ce clade est le taxon frère d'un clade composé des espèces Cearadactylus atrox, Brasileodactylus araripensis, Ludodactylus sibbicki, ainsi que des familles Ornithocheiridae et Anhangueridae, qui sont toutes membres du groupe Pteranodontoidea[7]. Bien que Myers ait trouvé qu'Aetodactylus était étroitement lié à Cimoliopterus, les différences dans la morphologie et l'orientation des mâchoires et l'espacement des alvéoles indiquent qu'elles sont distinctes les unes des autres. En raison des similitudes dans la forme de la mâchoire ainsi que de la dentition de C. dunni et de C. cuvieri, et des différences claires d'Aetodactylus dans ces caractéristiques, il est peu probable que Cimoliopterus soit un groupe paraphylétique (non naturel) selon Myers[7].
En 2018, une analyse phylogénétique menée par le paléontologue américain Nicholas Longrich et ses collègues a également récupéré Aetodactylus et Cimoliopterus en tant que taxons frères, renforçant leur relation étroite[41]. En 2019, la paléontologue britannique Megan Jacobs et ses collègues ont effectué une analyse phylogénétique dans laquelle ils ont placé C. cuvieri et C. dunni au sein de la famille des Ornithocheiridae, en tant que taxon frère de Camposipterus nasutus. À leur tour, leur clade forme une polytomie (un groupe non résolu) avec les espèces Camposapterus colorhinus et C. sedgwickii[37]. Cette relation étroite entre C. cuvieri et Camposipterus a déjà été retrouvée par Hooley en 1914 et par Unwin en 2001, bien qu'en tant qu'espèces d'Ornithocheirus et d'Anhanguera, respectivement[2] - [19]. Toujours en 2019, le paléontologue brésilien Borja Holgado et ses collègues ont récupéré un clade constitué des Cimoliopterus et des Anhangueria, ce clade étant défini par un palais qui est dévié vers l'arrière. Ornithocheirus a été récupéré en tant que taxon frère de ce clade, et à son tour, tous ont été placés dans le groupe Ornithocheirae[42].

Plus tard en 2019, Pêgas et ses collègues ont redécrit l'espèce Ornithocheirus wiedenrothi sous le nom de Targaryendraco et ont nommé le nouveau groupe Targaryendraconia, dans lequel Cimoliopterus était inclus. Certaines des caractéristiques les plus notables de ce groupe incluent l'élancement des mâchoires, l'extrémité avant de la symphyse dentaire étant particulièrement comprimée latéralement par rapport aux autres lanceodontiens (le groupe plus large auquel ils appartiennent), sa largeur totale égale à environ trois fois la largeur de l'alvéole dentaire, ainsi que le contour latéral subparallèle des marges de la moitié antérieure des deux mâchoires en vue occlusale (vue de dessous les dents)[30]. Pêgas et ses collègues ont découvert que Cimoliopterus, Camposipterus et Aetodactylus forment un groupe qu'ils ont nommé Cimoliopteridae. Ce groupe est soutenu par plusieurs synapomorphies (traits dérivés partagés). Cimoliopterus et Camposipterus partagent la caractéristique d'avoir les trois premières paires d'alvéoles dentaires supérieures plus proches l'une de l'autre que les postérieures, avec la troisième plus proche de la seconde que de la quatrième, le même motif est observé chez Aetodactylus mais dans sa mâchoire inférieure au lieu de la supérieure. D'autres caractéristiques communes entre Cimoliopterus et Camposipterus incluent la présence d'une crête palatine qui s'étend vers l'avant jusqu'au niveau entre la troisième et la deuxième paire d'alvéoles dentaires ainsi que l'emplacement de la partie la plus large de la partie avant du museau, qui est à la troisième paire d'alvéoles dentaires. Le cladogramme de l'analyse phylogénétique de Pêgas et ses collègues est présenté ci-dessous à droite, montrant la position des Cimoliopterus au sein des Cimoliopteridae, tandis que les autres targaryendraconiens, Aussiedraco, Barbosania et Targaryendraco, étaient regroupés en Targaryendraconidae[30].
|
Cladogramme 1 : Jacobs and colleagues, 2019.
|
Cladogramme 2 : Pêgas and colleagues, 2019.
|
Évolution

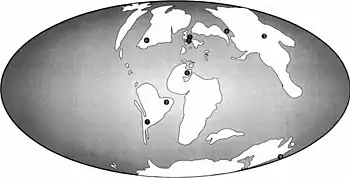
Unwin a déclaré en 2001 que le Cambridge Greensand et ce qui est maintenant le sous-groupe Grey Chalk ont fourni certains des plus jeunes enregistrements d'ornithocheiridés, A. cuvieri étant le plus jeune membre connu du groupe[2]. En raison de la crête prémaxillaire positionnée plus vers l'arrière des Cimoliopterus, contrairement à celles observées chez les anhangueridés, Rodrigues et Kellner ont suggéré en 2013 que leurs crêtes auraient pu évoluer indépendamment les unes des autres[38]. En 2015, Myers a déclaré que la découverte de C. dunni en Amérique du Nord a étendu la distribution du genre Cimoliopterus, montrant que les faunes de ptérosaures d'Europe et d'Amérique du Nord étaient similaires au milieu du Crétacé malgré l'élargissement continu de l'océan Atlantique Nord. Le fait que les ptérosaures aient conservé des affinités paléobiogéographiques (distribution géographique des groupes d'animaux préhistoriques) au milieu du Crétacé a également été confirmé par d'autres ptérosaures apparentés identifiés en Amérique du Nord, en Europe et en Afrique du Nord, tels que Coloborhynchus et Uktenadactylus[7] .
Myers a expliqué dans un communiqué de presse que la population ancestrale de C. dunni et C. cuvieri était capable de se déplacer entre l'Amérique du Nord et l'Angleterre jusqu'à il y a environ 94 millions d'années, car la similitude entre les deux espèces indiquait qu'il y avait eu peu de temps entre leur divergence. Lorsque l'Atlantique a ouvert le supercontinent Pangée, les populations d'animaux se sont isolées les unes des autres et ont divergé au cours de l'évolution, mais ce schéma aurait été plus compliqué avec les ptérosaures car ils pouvaient voler à travers les plans d'eau. Myers n'a pas suggéré la présence d'un pont terrestre entre les continents, mais que le niveau de la mer a fluctué avec le temps, et les ptérosaures auraient pu sauter d'île en île entre les masses terrestres qui ont émergé entre eux. Cependant, il restait encore un mystère pourquoi aucun lien étroit n'avait été identifié entre les ptérosaures d'Amérique du Nord et du Sud, indiquant qu'il aurait pu y avoir un obstacle à leur dispersion[31] .
En 2019, Pêgas et ses collègues ont convenu avec Rodrigues et Kellner que la crête de Cimoliopterus avait évolué indépendamment de celles des anhangueriens, car les taxons targaryendraconiens plus proches, tels que Barbosania, Camposipterus et Aetodactylus, ne possédaient pas de crêtes. Selon leur analyse, puisque Cimoliopterus était plus étroitement lié à Targaryendraco (d'âge Hauterivien), Aussiedraco (d'âge Albien) et Barbosania (également Albien) qu'aux anhangueriens, cela aiderait à combler un vide temporel qui entraînerait autrement une lignée fantôme menant à Cimoliopterus remontant à l'âge Valanginien. Pêgas et ses collègues pensaient que la légère expansion latérale des mâchoires d'Aetodactylus et de Cimoliopterus différait de l'état semblable à une rosette observé dans les mâchoires des anhangueriens, et qu'il n'était pas clair si l'état du premier groupe était un stade précurseur de celui du dernier. , ce problème nécessitant une enquête plus approfondie. Ils ont conclu que la reconnaissance du clade Targaryendraconia montrait que la lignée des Cimoliopterus était un groupe frère diversifié et cosmopolite d'Anhanguera, les deux ayant une distribution stratigraphique similaire[30].
Paléobiologie
Alimentation
.jpg.webp)
En 1987, Wellnhofer a déclaré que si les crêtes des ptérosaures tels que Pteranodon et Dsungaripterus étaient probablement utilisées comme gouvernails aérodynamiques ou comme zones d'attache musculaire, les crêtes situées à l'avant des mâchoires de Tropeognathus et de ses proches ne pouvaient pas servir à cet effet. Il a proposé à la place que ces crêtes auraient pu fonctionner comme la quille d'un bateau, pour stabiliser les mâchoires dans l'eau tout en écumant et en attrapant des poissons avec les mâchoires immergées à grande vitesse. La forme de la quille réduirait la résistance à l'écoulement, maintenant la tête dans une position stable, et aurait donc une fonction hydrodynamique plutôt qu'aérodynamique[22]. En 1991, il a précisé qu'ils auraient pêché en volant juste au-dessus de l'eau, en poussant rapidement la longue tête vers l'avant, en la plongeant dans l'eau et en saisissant des proies avec leurs mâchoires pointues[24].
Le paléontologue néerlandais André J. Veldmeijer et ses collègues ont souligné en 2006 qu'en dehors des ornithocheiridés généralement trouvés dans les dépôts associés à l'eau, leurs dents imbriquées soutenaient également la piscivorie (manger du poisson), étant construite pour harponner des proies glissantes plutôt que de couper ou de taillader. Les longues dents de devant incurvées conviendraient à la capture de proies glissantes telles que les poissons, et les dents plus petites à l'arrière des mâchoires garantissaient que la nourriture était retenue et transportée dans la gorge. La position frontale des yeux et certaines adaptations du cerveau peuvent avoir été liées à l'évaluation précise de la position d'un poisson avant d'entrer dans l'eau. Ils ont également souligné que les différences de position de la crête, la taille de la crête palatine et la présence ou l'absence d'une expansion antérieure de la mâchoire rendaient difficile de croire qu'ils obtenaient tous de la nourriture de la même manière, mais que cela n'excluait pas certains chevauchements[43].

Veldmeijer et ses collègues ont noté que puisque les corps de ces ptérosaures étaient petits, ils n'auraient pas eu d'espace pour les gros poissons, et ces poissons peuvent également avoir altéré leur capacité de vol, comme c'est le cas pour les chauves-souris. Ils peuvent avoir chassé de petits poissons ou les avoir pré-digérés avant de les avaler (puisque leurs dents n'étaient pas adaptées à la mastication), mais la deuxième option aurait nécessité des joues ou des sacs gulaire pour garder les proies à l'intérieur de la bouche ; ce dernier a été signalé chez certains ptérosaures. Ils ont exclu l'écrémage comme méthode d'alimentation, car ils pensaient que cela poserait des problèmes de stabilité en vol si un bout d'aile touchait la surface de l'eau. Les auteurs ont soutenu l'hypothèse de capture de poisson de Wellnhofer en testant un modèle géométrique basé sur le crâne et le cou d'Anhanguera, et ont suggéré qu'il se serait approché de l'eau avec sa tête parallèle à la surface, l'aurait tournée vers le bas tout en étendant le cou pour atteindre le poisson en mouvement le plus lent, tandis que la crête aidait à maintenir la stabilité en contrecarrant la force de l'eau. Ils ont conclu que les crêtes prémaxillaires étaient probablement pour minimiser l'attrition de l'eau, et donc pas pour la parade nuptiale. Ils n'ont pas pu déterminer quelles étaient les conséquences d'Anhanguera ayant une crête placée plus vers l'arrière que Coloborhynchus et Ornithocheirus[43].
Witton a déclaré en 2012 que les ornithocheiridés montraient des adaptations pour se nourrir sans atterrir, avec des dents de taille variable qui dépassaient de la mâchoire sous de nombreux angles. Utiliser uniquement les dents de devant pour attraper une proie a permis d'éloigner le corps de la surface de l'eau, minimisant ainsi le risque de s'écraser[44]. En 2013, Witton a noté que l'hypothèse d'alimentation écrémée pour les ornithocheiridés avait été remise en question, mais que l'alimentation par immersion (comme on le voit chez les sternes et les frégates) était soutenue par diverses caractéristiques, comme leurs museaux allongés, bien adaptés pour atteindre les animaux nageurs, ainsi que leur arrangement de dents en « attrape-poisson ». Witton doutait de l'idée que les crêtes prémaxillaires aient été utilisées pour stabiliser l'extrémité des mâchoires lorsqu'elles étaient plongées dans l'eau pour se nourrir, car certains ornithocheiridés manquaient complètement de telles crêtes, et les mangeurs aériens modernes peuvent obtenir la même chose sans structures similaires. La diversité de la morphologie des dents chez les ornithocheiridés indique que des méthodes d'alimentation autres que l'alimentation par immersion étaient également utilisées par les membres du groupe, comme chercher de la nourriture à la surface de l'eau ou plonger à faible profondeur sous la surface[1]. Au lieu d'être utilisé pour la stabilisation pendant l'écrémage ou le vol, Witton considérait que les crêtes des ptérosaures étaient des structures de parade, car les crêtes auraient été sexuellement dimorphes chez certaines espèces[45].
Locomotion

Witton a résumé les idées sur les capacités locomotrices des ornithocheiridés en 2013 et a déclaré que des caractéristiques telles que de petites pattes mais de grandes ailes, de faibles masses corporelles pour leurs envergures, des ailes avec des rapports d'aspect élevés et une charge alaire réduite indiquent qu'ils préféraient la vie dans les airs et étaient adapté au vol plané sur de longue distance. Étant donné que leurs formes d'ailes étaient similaires à celles des oiseaux de mer océaniques, ils étaient probablement mieux adaptés au vol océanique qu'au vol au-dessus des zones terrestres, ce qui est soutenu par le fait que leurs fossiles se trouvent principalement dans les dépôts marins. Ils étaient peut-être capables de décoller de l'eau, ce qui serait aidé par leurs petits torses et leurs petites jambes. Leur locomotion terrestre était probablement limitée par leurs pattes courtes, et ils ont peut-être été limités à des marches traînantes plus lentes (de sorte que les longs membres antérieurs ne dépassent pas les membres postérieurs) et à des sauts plus rapides, ce qui indique qu'ils ont peut-être passé un temps limité au sol. Comme cela a été supposé pour les ptérosaures en général, ils ont probablement pris leur envol en se lançant en quadrupède, et ils étaient peut-être capables de nager assez bien pour pouvoir décoller des surfaces d'eau[1].
En 2017, le biologiste britannique Colin Palmer a utilisé des tomodensitométries d'os d'aile de ptérosaure ornithocheiridé, y compris la phalange d'aile qui avait été attribuée à C. cuvieri, pour reconstruire le longeron d'aile (le principal élément structurel) des membranes d'aile de ptérosaure. Il a utilisé la résistance à la flexion du longeron d'aile d'un modèle structurel avec une envergure d'aile de 6 mètres sur la base des tomodensitométres pour déduire la tension probable de la membrane de l'aile nécessaire pour supprimer le flottement aéroélastique (instabilité causée par la résistance de l'air) à des vitesses de vol élevées et en minimisant le gonflement, empêchant ainsi la défaillance structurelle de la membrane sous les charges de vol. Les trois estimations étaient d'une ampleur similaire, ce qui implique que la membrane aurait un matériau à module élevé (module de Young, élasticité en traction), ce qui soutient l'idée que la couche d'aktinofibrilles (fibres structurelles à l'intérieur des ailes des ptérosaures) à l'extérieur de la membrane qui a renforcé les ailes des ptérosaures étaient en matière kératinique (la même matière dans les cheveux et les ongles humains) et différaient grandement des membranes des chauves-souris[46].
Paléoenvironnement
L'holotype de C. cuvieri a été trouvé dans le Kent, en Angleterre, collecté dans des craies et des marnes du sous-groupe de la craie grise, qui date des âges Cénomanien-Turonien du Crétacé supérieur, bien que la position stratigraphique précise du spécimen soit inconnue. Le sous-groupe de Grey Chalk a également été historiquement connu sous le nom de formation de Chalk inférieur ou Chalk Marl, et est maintenant considéré comme un membre du groupe de Chalk, lui-même également connu sous le nom de formation de Chalk[7] - [9] - [47]. Le sous-groupe de la craie grise est constitué de dépôts marins peu profonds, composés de craie argileuse ou marneuse sans silex, et a été géologiquement daté à partir de zones biostratigraphiques définies par la présence des ammonites Mantelliceras mantelli et Calycoceras guerangeri (fossiles index utilisés pour dater les couches)[31] - [47] - [48]. D'autres animaux connus du sous-groupe de la craie grise comprennent les ptérosaures tels que Lonchodraco et de nombreuses espèces douteuses, et les dinosaures comme le nodosauridé indéterminé Acanthopholis et l'hadrosauroide « Iguanodon » hilli[47].
L'holotype de C. dunni a été trouvé dans la formation de Britton du centre-nord du Texas, qui remonte au stade cénomanien du début du Crétacé supérieur. La formation fait partie du groupe Eagle Ford du Crétacé supérieur, qui date du Cénomanien moyen à la fin du Turonien (il y a 96 à 90 millions d'années). Le spécimen a été conservé dans une couche de schiste marin gris avec des concrétions d'oxyde de fer et trouvé dans la zone d'ammonite Sciponoceras gracile, située dans la partie supérieure-moyenne de la Formation de Britton, qui date de la fin du Cénomanien, il y a environ 94 millions d'années. D'abondants restes fossiles d'ammonites et de crustacés sont contenus dans les schistes gris foncé dans lesquels C. dunni a été trouvé, ce qui est cohérent avec le fait qu'ils s'étaient déposés dans des environnements marins de plateau faible en énergie et mal oxygénés[7]. La zone dans laquelle il a été trouvé se trouvait au large de la voie maritime intérieure de l'Ouest peu profonde, qui couvrait la partie centrale des États-Unis et du Canada[31]. Les autres animaux de la formation de Britton comprennent des décapodes, des ammonites, des poissons (y compris des requins), des plésiosaures, des tortues et coniasaurs (en). Des coprolithes (fèces fossiles) attribués aux poissons sont également connus[49].
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Cimoliopterus » (voir la liste des auteurs).
- Witton 2013, p. 152–163.
- D. M. Unwin, « An overview of the pterosaur assemblage from the Cambridge Greensand (Cretaceous) of Eastern England », Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe, vol. 4, , p. 189–221 (DOI 10.1002/mmng.20010040112
 )
) - Rodrigues et Kellner 2013, p. 4–17.
- D. M. Martill, « The early history of pterosaur discovery in Great Britain », Geological Society, London, Special Publications, vol. 343, no 1, , p. 287–311 (DOI 10.1144/SP343.18, Bibcode 2010GSLSP.343..287M, S2CID 130116778)
- J. S. Bowerbank, « On the pterodactyles of the Chalk Formation », Proceedings of the Zoological Society of London, vol. 19, , p. 14–20 (DOI 10.1111/j.1096-3642.1851.tb01125.x, lire en ligne)
- Witton 2013, p. 98.
- T. S. Myers, « First North American occurrence of the toothed pteranodontoid pterosaur Cimoliopterus », Journal of Vertebrate Paleontology, vol. 35, no 6, , p. 1–9 (DOI 10.1080/02724634.2015.1014904, S2CID 86099117, lire en ligne)
- J. A. Jobling, The Helm Dictionary of Scientific Bird Names, London, Christopher Helm, (ISBN 978-1-4081-2501-4, lire en ligne), p. 125
- Rodrigues et Kellner 2013, p. 37–45.
- R. Owen, A History of British Fossil Reptiles, vol. 1, London, Cassell & Company Limited, 1849–1884, 242–258 p. (DOI 10.5962/bhl.title.7529, lire en ligne)
- C. Palmer, « Inferring the properties of the pterosaur wing membrane », Geological Society, London, Special Publications, vol. 455, no 1, , p. 57–68 (DOI 10.1144/SP455.4, Bibcode 2018GSLSP.455...57P, S2CID 132668858)
- R. Owen, Geology and Inhabitants of the Ancient World, vol. 8, London, Crystal Palace library, , 5-7, 11-13 (ISBN 978-1-166-91304-5, lire en ligne)
- M. Witton, « Pterosaur – information about the Crystal Palace statues », sur cpdinosaurs.org, (consulté le )
- F. Knoll et R. López-Antoñanzas, « Lasuén's pterodactyl: An early use of a pterosaur in plastic arts », Comptes Rendus Palevol, vol. 9, no 5, , p. 245–254 (DOI 10.1016/j.crpv.2010.07.001)
- M. Witton, « The science of the Crystal Palace Dinosaurs, part 2: Teleosaurus, pterosaurs and Mosasaurus », sur Mark Witton.com Blog, (consulté le )
- H. G. Seeley, Index to the Fossil Remains of Aves, Ornithosauria, and Reptilia, from the Secondary System of Strata Arranged in the Woodwardian Museum of the University of Cambridge, Cambridge, Deighton, Bell, and co., (DOI 10.5962/bhl.title.159380 , lire en ligne), p. 16
- H. G. Seeley, The Ornithosauria: an Elementary Study of the Bones of Pterodactyls, Made from Fossil Remains Found in the Cambridge Upper Greensand, and Arranged in the Woodwardian Museum of the University of Cambridge, Cambridge, Deighton, Bell, and Co., (lire en ligne), p. 113
- R. Owen, A Monograph on the Fossil Reptilia of the Mesozoic Formations, vol. 1, London, The Palæontographical Society, , 6–8 p. (DOI 10.1017/CBO9781316151099.003)
- R. W. Hooley, « LXI.— On the ornithosaurian genus Ornithocheirus, with a review of the specimens from the Cambridge Greensand in the Sedgwick Museum, Cambridge », Annals and Magazine of Natural History, vol. 13, no 78, , p. 529–557 (DOI 10.1080/00222931408693521, lire en ligne)
- (de) G. von Arthaber, « Über Entwicklung, Ausbildung und Absterben der Flugsaurier », Paläontologische Zeitschrift, vol. 4, , p. 1–47 (DOI 10.1007/BF03041557, S2CID 131644821, lire en ligne)
- (de) F. B. Nopcsa, « Bemerkungen und ergänzungen zu G. V. ARTHABERs Arbeit über Entwicklung und Absterben der Pterosaurier », Paläontologische Zeitschrift, vol. 6, no 1, , p. 80–91 (DOI 10.1007/BF03041533, S2CID 140187167)
- P. Wellnhofer, « New crested pterosaurs from the Lower Cretaceous of Brazil », Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie, vol. 27, , p. 175–186 (lire en ligne)
- D. A. Campos et A. W. A. Kellner, « Panorama of the flying reptiles study in Brazil and South America », Anais da Academia Brasileira de Ciencias, vol. 57, no 4, , p. 459–465 (lire en ligne)
- Wellnhofer 1991.
- S. C. Bennett, « The ontogeny of Pteranodon and other pterosaurs », Paleobiology, vol. 19, no 1, , p. 92–106 (ISSN 0094-8373, DOI 10.1017/S0094837300012331, JSTOR 2400773, lire en ligne)
- N. N. Bakhurina et D. M. Unwin, « A survey of pterosaurs from the Jurassic and Cretaceous of the former Soviet Union and Mongolia », Historical Biology, vol. 10, no 3, , p. 197–245 (DOI 10.1080/10292389509380522)
- D. M. Unwin, J. Lü et N. N. Bakhurina, « On the systematic and stratigraphic significance of pterosaurs from the Lower Cretaceous Yixian Formation (Jehol Group) of Liaoning, China », Mitteilungen aus dem Museum für Naturkunde in Berlin, Geowissenschaftliche Reihe, vol. 3, , p. 181–206 (DOI 10.1002/mmng.20000030109 )
- A. W. A. Kellner, T. Rodrigues et F. R. Costa, « Short note on a pteranodontoid pterosaur (Pterodactyloidea) from western Queensland, Australia », Anais da Academia Brasileira de Ciências, vol. 83, no 1, , p. 301–308 (PMID 21437387, DOI 10.1590/S0001-37652011000100018 )
- Rodrigues et Kellner 2013, p. 70.
- R. V. Pêgas, B. Holgado et M.E.C. Leal, « On Targaryendraco wiedenrothi gen. nov. (Pterodactyloidea, Pteranodontoidea, Lanceodontia) and recognition of a new cosmopolitan lineage of Cretaceous toothed pterodactyloids », Historical Biology, vol. 33, no 8, , p. 1–15 (DOI 10.1080/08912963.2019.1690482, S2CID 209595986)
- M. Allen, « North America's newest pterosaur is a Texan — and flying reptile's closest cousin is English », SMU Research, (lire en ligne, consulté le )
- J.S. Bowerbank, « On a new species of pterodactyl found in the Upper Chalk of Kent (Pterodactylus giganteus) », Quarterly Journal of the Geological Society of London, vol. 2, , p. 7–9 (DOI 10.1144/gsl.jgs.1846.002.01-02.05, S2CID 129389179, lire en ligne)
- R. Owen, « On a new species of pterodactyle (Pterodactylus compressirostris, Owen) from the Chalk; with some remarks on the nomenclature of the previously described species », Proceedings of the Zoological Society of London, vol. 19, no 1, , p. 21–34 (DOI 10.1111/j.1096-3642.1851.tb01126.x, lire en ligne)
- Witton 2013, p. 51–52.
- Rodrigues et Kellner 2013, p. 54–64.
- B. Holgado et R. V. Pêgas, « A taxonomic and phylogenetic review of the anhanguerid pterosaur group Coloborhynchinae and the new clade Tropeognathinae », Acta Palaeontologica Polonica, vol. 65, no 4, , p. 743–761 (DOI 10.4202/app.00751.2020 )
- Jacobs, M. L., Martill, D. M., Ibrahim, N. et Longrich, N., « A new species of Coloborhynchus (Pterosauria, Ornithocheiridae) from the mid-Cretaceous of North Africa », Cretaceous Research, vol. 95, , p. 77–88 (DOI 10.1016/j.cretres.2018.10.018, S2CID 134439172, lire en ligne)
- Rodrigues et Kellner 2013, p. 72–88.
- X. Wang, T. Rodrigues, S. Jiang, X. Cheng et A. W. A. Kellner, « An Early Cretaceous pterosaur with an unusual mandibular crest from China and a potential novel feeding strategy », Scientific Reports, vol. 4, , Article number 6329 (PMID 25210867, PMCID 5385874, DOI 10.1038/srep06329)
- B. Andres et T. S. Myers, « Lone star pterosaurs », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 103, nos 3–4, , p. 383–398 (DOI 10.1017/S1755691013000303, S2CID 84617119)
- N. R. Longrich, D. M. Martill, B. Andres et D. Penny, « Late Maastrichtian pterosaurs from North Africa and mass extinction of Pterosauria at the Cretaceous-Paleogene boundary », PLOS Biology, vol. 16, no 3, , p. 23–24 (PMID 29534059, PMCID 5849296, DOI 10.1371/journal.pbio.2001663)
- B. Holgado, R. V. Pêgas, J. I. Canudo, J. Fortuny, T. Rodrigues, J. Company et A. W. A. Kellner, « On a new crested pterodactyloid from the Early Cretaceous of the Iberian Peninsula and the radiation of the clade Anhangueria », Scientific Reports, vol. 9, no 1, , p. 1–10 (PMID 30894614, PMCID 6426928, DOI 10.1038/s41598-019-41280-4 , Bibcode 2019NatSR...9.4940H)
- A. J. Veldmeijer, M. Signore et E. Bucci, Predation in Organisms: A Distinct Phenomenon, , 295–308 p. (ISBN 978-3-540-46044-2, DOI 10.1007/978-3-540-46046-6_13), « Predator-prey interaction of Brazilian Cretaceous toothed pterosaurs: a case example »
- A. J. Veldmeijer, M. Witton et I. Nieuwland, Pterosaurs: Flying Contemporaries of the Dinosaurs, Leiden, Sidestone Press, , 81–83, 98 (ISBN 978-90-8890-093-8)
- Witton 2013, p. 79–82.
- C. Palmer, « Inferring the properties of the pterosaur wing membrane », Geological Society, London, Special Publications, vol. 455, no 1, , p. 57–68 (DOI 10.1144/SP455.4, Bibcode 2018GSLSP.455...57P, S2CID 132668858)
- Z. Csiki-Sava, E. Buffetaut, A. Ősi, X. Pereda-Suberbiola et S. L. Brusatte, « Island life in the Cretaceous – faunal composition, biogeography, evolution, and extinction of land-living vertebrates on the Late Cretaceous European archipelago », ZooKeys, no 469, , p. 14–15 (PMID 25610343, PMCID 4296572, DOI 10.3897/zookeys.469.8439 )
- P. M. Hopson, A stratigraphical framework for the Upper Cretaceous Chalk of England and Scotland with statements on the Chalk of Northern Ireland and the UK offshore sector, British Geological Survey, , 8–15 p. (ISBN 978-0-85272-517-7, lire en ligne)
- V. Friedman, Paleoecology and Depositional Environment of the lower Eagle Ford Group in North Central Texas (thèse), The University of Texas at Dallas, , 71–73 p. (lire en ligne)
Ouvrages cités
- P. Wellnhofer, The Illustrated Encyclopedia of Pterosaurs, New York, Crescent Books, (ISBN 978-0-517-03701-0)
- M. P. Witton, Pterosaurs: Natural History, Evolution, Anatomy, Princeton and Oxford, Princeton University Press, , 1st éd. (ISBN 978-0-691-15061-1)
- T. Rodrigues et A. Kellner, « Taxonomic review of the Ornithocheirus complex (Pterosauria) from the Cretaceous of England », ZooKeys, no 308, , p. 1–112 (PMID 23794925, PMCID 3689139, DOI 10.3897/zookeys.308.5559 )
Liens externes
- Ressource relative au vivant :