Règle de Bergmann
La règle de Bergmann est, en zoologie, un principe observé au sein d'un clade, qui corrèle la température de l'environnement avec le ratio surface/volume du corps chez les animaux endothermes, ceux dont la température intérieure est indépendante du milieu.
Historique
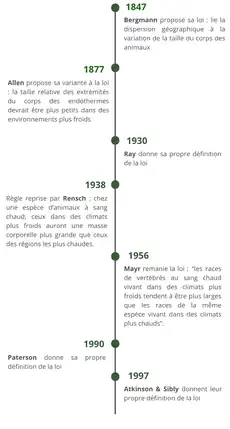
La loi de Bergmann est une règle éco-géographique proposée par Carl Bergmann en 1847. Cette règle lie la dispersion géographique des animaux à la variation de la taille du corps des organismes. Carl Bergmann écrit la définition suivante: “ Si deux espèces d’un même genre ne diffèrent que par leur taille alors la plus petite espèce vit sous un climat plus chaud et plus au Sud que l’espèce plus grosse vivant au Nord”[1] - [2].
En 1938, la règle est reprise par Rensch. Il définit la règle comme s’appliquant à “un complexe de race d’animaux au sang chaud tel que les races vivant dans les régions plus froides sont généralement plus grandes que celles vivant dans les régions plus chaudes”. Cette nouvelle définition est devenue la définition de référence de la règle de Bergmann. En 1956, Mayr remanie lui aussi la règle de Bergmann : “les races de Vertébrés au sang chaud vivant dans des climats plus froids tendent à être plus grandes que les races de la même espèce vivant dans des climats plus chauds”. De nombreux autres auteurs comme Ray (1930), Paterson (1990) ou Atkinson & Sibly en 1997 ont eux aussi tenté d’établir une définition de la règle de Bergmann[2].
Aujourd’hui, la communauté scientifique cherche à définir les conditions d’application (e.g. phylogénie) et les mécanismes derrière la règle de Bergmann. De plus, les définitions modernes cherchent à étendre la règle (initialement appliquée aux endothermes : mammifères, oiseaux…) à tous les animaux y compris des ectothermes.
Carl Bergmann relie la distribution géographique d’endothermes avec leur taille et leur bilan thermique sans préciser les mécanismes en jeu. La majorité des définitions de la règle ne comprennent pas d'explication du phénomène[2]. La corrélation de la taille et de la température suggère que les mécanismes de thermorégulation et de rétention de la chaleur sont des facteurs conduisant à l’évolution de la taille[3]. En effet, les corps trapus sont favorisés dans les environnements froids car la faible surface de contact avec l’extérieur limite la dissipation de la chaleur. Ce qui signifie que le rapport surface / volume est impliqué dans la thermorégulation. La taille pourrait ainsi être considérée comme une adaptation thermorégulatrice pour répondre aux différentes conditions climatiques. Les endothermes auraient un rapport surface / volume faible à des latitudes et/ou des altitudes plus élevées[4] - [5].
Pour Rensch et Mayr, la règle de Bergmann est empirique. De plus, Mayr considère la règle de Bergmann comme étant liée à la sélection naturelle[6]. Toutefois, la masse corporelle peut aussi être influencée par les fluctuations saisonnières et journalières et par les conditions de reproduction[7]. Selon Mayr, si 50% des espèces étudiées sont conformes à cette loi, la validité de celle-ci ne devrait pas être remise en question.
En 1936, Rensch a conclu que la majorité des mammifères Nord-Américains et Européens se conforment à cette loi, cependant, Scholander (en 1955) a remis en question les choix de caractéristiques prises en compte par Rensch dans ses recherches.
D’autres caractéristiques ont été utilisées comme substituts à la masse corporelle, tels que la taille des ailes ou encore la taille des dents. Néanmoins, ces caractéristiques ne reflètent pas bien la masse corporelle car toutes deux peuvent être influencées par des paramètres différents[7]. De plus, la définition la plus admise actuellement considère une variation de taille selon la latitude et non plus uniquement en termes d’environnements chauds ou froids[8]. Le climat et la latitude sont fortement corrélés même si cela dépend de l’échelle considérée[2].
Généralités: Mammifères, Oiseaux
La loi nommée par le biologiste allemand Carl Bergmann, s'applique aux endothermes, des animaux thermorégulateurs produisant eux-mêmes leur chaleur interne. Leur température interne reste donc stable, malgré les conditions externes, par des mécanismes d'homéostasie. Ainsi, les oiseaux, les mammifères, les amphibiens et les tortues des climats froids tendent à avoir une masse corporelle plus importante que ceux des climats chauds. Par exemple les chevreuils à queue blanche sont plus grands au Canada que dans les Keys de Floride. Les cas des sangliers, des manchots ou des ours sont également fréquemment cités.
F. Delpech, qui cite le cas du Cervus simplicidens, du renne, de la hyène des cavernes et d'autres animaux, note que les températures froides amènent un environnement steppique, des tailles d'individus plus grandes et des populations moins importantes ; tandis que les températures plus clémentes correspondent à un habitat forestier, des tailles d'individus plus petites et des populations plus importantes. Elle constate que « la mise en place de la forêt, qu'il s’agisse de la taïga ou de la forêt caducifoliée, entraîne toujours une baisse de la biomasse des ongulés », et que la réduction importante de taille des carnivores est en corrélation avec la raréfaction du gibier.
Oiseaux
Selon plusieurs analyses, 72 à 76 % des espèces d’oiseaux suivraient cette loi. Ces proportions proviennent d'études sur des espèces issues de différents ordres et familles[7] - [6]. Ces études ont observé une conformité significativement plus importante de la loi chez les espèces sédentaires que chez les espèces migratrices. En effet, les espèces passant l’hiver dans des régions plus chaudes sont potentiellement moins sujettes à la sélection naturelle de ce trait (taille de l’organisme)[7].
Exemple de chouette du genre Tyto (famille des Tytonidae)[4]:

L’étude menée par la chercheuse Andréa Romano et son équipe a mis en avant la variation de la biomasse de plusieurs espèces de chouettes à travers différents climats continentaux. Leurs travaux montrent que la distribution de la taille des ailes suit un gradient de température. Ainsi la taille des ailes est plus grande chez les individus vivant dans des milieux où la température moyenne annuelle est la plus basse. Cette caractéristique répond à l’hypothèse de la conservation de la chaleur. La biomasse serait donc une caractéristique convergente répondant aux mécanismes de thermorégulation et donc aux conditions thermiques environnementales des groupes d’espèces d’oiseaux considérés dans l’étude. Le modèle évolutif des individus trapus dans les climats froids semble être principalement conduit par la variation de températures latitudinale que par la variation altitudinale. L’amplitude de la variation de taille des ailes intraspécifique est plus importante avec la latitude qu’avec l’altitude. De plus, la taille des individus est mieux prédite par la variation de température minimum que par la variation de température maximum annuelle. Ceci peut s’expliquer par le fait que les chouettes sont des oiseaux nocturnes, connus pour être sensibles aux hivers extrêmement froids. De plus, les oiseaux nocturnes ne sont pas particulièrement exposés aux températures trop élevées. En effet, ils sont généralement actifs dans les moments les plus froids de la journée.
Mammifères
De même, selon les études, 65 à 82% des espèces de mammifères présenteraient une corrélation positive entre la biomasse des individus et la latitude ou température[9]. Cependant, d’autres études montrent une plus faible tendance à suivre la loi de Bergmann chez les mammifères ayant une masse corporelle inférieure à 500 g. Cela peut s'expliquer par le fait que beaucoup d’espèces de micromammifères vivent dans des terriers donc dans des microclimats favorables. De plus, plusieurs espèces de rongeurs hibernent durant les mois d’hiver. Ainsi, les écologues distinguent des espèces aux stratégies de tolérance ou d’évitement face à la contrainte. La loi de Bergmann traduirait une forme de tolérance via une adaptation anatomique, tandis que l’entrée en dormance, une forme d’évitement[7].
Néanmoins, cette règle comporte de nombreuses exceptions connues. Bien que formulée au départ pour des types d'animaux considérés comme une espèce d'un même genre, la règle est aujourd'hui peut-être appliquée aux sous-espèces.
Exemple et contre-exemple du Renard des savanes[10]:

La variation de la masse corporelle du renard des savanes Cerdocyon thous a été analysée sur un gradient de latitude de son aire de distribution en Amérique du Sud. Le but étant de déterminer si ce mammifère suit la règle de Bergmann au nord et au sud de l'équateur.
Cette analyse révèle deux tendances distinctes. Les individus au sud de l'équateur montrent une augmentation significative de la taille avec l'augmentation de la latitude. Cependant, les populations au nord de l'équateur suivent une tendance opposée; une diminution significative de la masse corporelle avec l'augmentation de la latitude. La règle de Bergmann est donc respectée pour les individus au sud de l’équateur, mais non pas pour ceux au nord de l’équateur. En cherchant à comprendre cette observation, les chercheurs ont relevé la présence de deux sous-espèces dans le nord de l’équateur, C. t. aquilus et C. t. thous, aux masses corporelles différentes. Cette déviation par rapport à la règle de Bergmann dans les populations du Nord serait donc influencée par des facteurs historiques, l'adaptation locale, la sélection naturelle et la dérive génétique.
Extension de la règle de Bergmann aux ectothermes
Historiquement, la loi de Bergmann a été formulée exclusivement pour les organismes endothermes. Cependant, de nouvelles études ont également permis de relier cette loi aux Vertébrés ectothermes, ainsi que chez de nombreux taxons d’Invertébrés[7]. Il y a une vingtaine d’années, Ray a montré que la règle était soutenue pour 75% des espèces de ectothermes[2]. En revanche, aujourd’hui on ne sait pas si ce chiffre est toujours valable.
Les Chéloniens et les Squamates
Ashton et Feldman[8] se sont intéressés à la relation entre latitude/altitude et la température pour deux groupes de reptiles situés dans l’hémisphère Nord : les Chéloniens (tortues) et les Squamates (lézards et serpents). Ces organismes sont étudiés car ils ne possèdent pas de faibles restrictions latitudinales et altitudinales, contrairement aux Crocodiliens et aux Sphénodons.
Dans l’hémisphère Nord, la majorité des Chéloniens suivent la règle de Bergmann contrairement aux Squamates qui suivent l’inverse de la règle.
Il a été démontré que différents mécanismes sont susceptibles d’intervenir dans la règle de Bergmann chez les ectothermes:
- La saisonnalité : les individus les plus gros sont susceptibles de mieux résister à une période de famine que les plus petits individus grâce à leur capacité supérieure de stockage de la graisse. Les périodes de disette sont plus fréquentes en hautes latitudes, dues à la forte saisonnalité des ressources. Cela serait vrai chez les Chéloniens au contraire des Squamates qui suivent l’inverse de la règle Bergmann.
- La compétition et la sélection, pour le maintien d’une température corporelle préférentielle, peuvent être différentes selon les organismes. À des latitudes élevées, la taille corporelle est grande en raison d'une compétition détendue. Mais cela est différent chez les Squamates, qui pourraient évoluer vers des tailles corporelles plus petites lorsque la compétition se relâche.
- L’hypothèse de la conservation de la chaleur chez les Chéloniens : dans les milieux plus froids, la coquille, le plan corporel dense et le faible rapport surface/volume entraînent un réchauffement et un refroidissement corporel lent. Cette inertie thermique présente des avantages dans la conservation de la chaleur. À l’inverse, dans des environnements frais, les Squamates auraient un ratio surface/volume plus important qui leur permettrait de se réchauffer plus vite aux hautes latitudes, et ainsi d’être actifs pendant une plus grande partie de la journée.
- La disponibilité des proies pourrait influencer directement la taille corporelle. Les organismes ayant une alimentation plus riche en énergie sont, à maturité sexuelle, plus grands que ceux ayant eu une alimentation plus faible en énergie (serpents)[8]. De même pour la qualité nutritionnelle des proies ou des végétaux consommés par les organismes[11].
Le crabe Minuca pugnax[11]
.jpeg.webp)
Minuca pugnax (en) est un excellent exemple pour examiner la règle de Bergmann. En effet, la distribution de chaque espèce de ce crabe peut s'étendre sur plusieurs degrés de latitude, et leur morphologie varie considérablement à l'intérieur de cette étendue. En moyenne, la largeur de leur carapace augmente de 0,5 mm à chaque degré de latitude. Les plus gros crabes sont trouvés dans la partie des États-Unis la plus éloignée de l’équateur (nord-est du Massachusetts), avec une largeur moyenne de carapace de 18,5 mm. Tandis que les plus petits se trouvent dans la partie des États-Unis située au niveau de l’équateur (en Géorgie), avec une largeur moyenne de carapace de 9,9 mm.
Parce que nos océans se réchauffent et que la taille du corps de M. pugnax (en) est inversement liée à la température, la taille du corps de cet animal sera fortement modifiée selon les latitudes à mesure que le climat continue de se réchauffer.
Critiques et ouvertures
Il n'existe pas encore de consensus scientifique quant à l'existence de cette règle[12]. Cela est dû en grande partie à la confusion qui règne quant au niveau taxonomique auquel elle est censée s’appliquer et à la multiplicité des facteurs en cause dans les mécanismes évolutifs et écologiques sous-jacents à cette règle[2].
Débat interspécifique et intraspécifique
Les interprétations des textes de Bergmann laisseraient penser que le modèle permet la comparaison des tailles d’individus au sein d’un niveau interspécifique. Cependant, la systématique et la phylogénie ayant énormément évolué depuis les travaux de Bergmann, les espèces étudiées pour son modèle n’appartiennent peut-être pas réellement à un même genre. De plus, de nombreux scientifiques, tels que Mayr, s'appuyaient sur la courbe de corrélation de Bergmann (augmentation de la taille avec la diminution de la température) pour décider l’appartenance ou non de deux populations à une même espèce. La règle de Bergmann était la preuve qu’au sein d’une même espèce, les populations pouvaient avoir des caractéristiques différentes selon les températures du milieu. Ainsi, les individus que Bergmann considérait comme appartenant à des espèces différentes pourraient aujourd’hui être perçus comme appartenant à des populations différentes d’une même espèce. La frontière entre l’inter- et l'intra spécifique est donc plutôt subjective pour ce modèle. L’application de la règle de Bergmann est donc encore très controversée[3].
Cependant, les recherches actuelles sur la règle de Bergmann privilégient l’intraspécifique[2]. En effet, l’application aux endothermes implique une forte évaluation de la loi de Bergmann sur le niveau intraspécifique. De plus, peu d’études visent à étudier cette loi de manière interspécifique chez les endothermes, et les seuls résultats ne permettent pas de souligner une corrélation de la taille avec la température[13].
Exemple d'exceptions infraspécifiques :
Une étude montre que seule une petite minorité d’espèces de Plethodon, un genre d’amphibiens suivraient la loi de Bergmann. De plus, une analyse comparative phylogénétique n'a trouvé aucune relation entre la taille corporelle et la température entre les espèces. Les résultats suggèrent alors que les relations entre taille corporelle et températures ne sont pas courantes chez les amphibiens. Il en a été conçu que la règle de Bergmann n'est généralement pas applicable à ces taxons[14].
En outre, la majorité des études montrant un non-suivi de la règle de Bergmann ont été faites sur des Invertébrés alors que celles montrant un suivi de la règle de Bergmann sur des Vertébrés[8].
Différents facteurs influençant la taille corporelle
Il est difficile de quantifier les relations entre la taille du corps et la géographie et cela peut expliquer les débats autour de la cause de la règle de Bergmann. De plus, d’autres mécanismes impactent la taille des organismes. La taille n’est pas uniquement impactée par les thermoclines[15].
Blackburn[2] a émis 6 hypothèses sur les mécanismes expliquant la variation spatiale de la taille:
- Hypothèse 1 : Les schémas spatiaux dans la taille du corps sont dus à une sélection de traits influençant indirectement la masse corporelle. Certains traits étant proches, une sélection d'un trait lié à la taille engendre une sélection indirecte du caractère taille.
- Hypothèse 2 : Les schémas spatiaux dans la taille du corps sont dus à une colonisation aléatoire de certains milieux par des ancêtres de grandes tailles et par la diversification de taxons qui en a suivi.
- Hypothèse 3 : Les espèces de grande taille auraient des capacités de dispersion supérieures à celles des espèces de petite taille dans les climats froids. Ainsi, les schémas spatiaux dans la taille du corps serait dus aux potentialités intrinsèques de l'espèce (capacité de propagation, aptitudes écologiques et évolutives).
- Hypothèse 4 : Conservation de la chaleur. La grande taille de certaines espèces leur permettrait d’occuper des milieux froids et/ou de hautes latitudes car leur taille leur permet de mieux conserver la chaleur grâce à un bon rapport surface/volume ou parce que ces espèces présentent des couches isolatrices plus épaisses.
- Hypothèse 5 : La taille des organismes serait liée au besoin de rester à une température corporelle peu élevée. Hypothèse privilégiée pour les habitats chauds et humides où la transpiration ne permet pas de se rafraîchir. Une des solutions est d’augmenter son ratio surface/volume et donc sa surface pour permettre une meilleure perte de chaleur.
- Hypothèse 6 : La disponibilité en ressources, montrerait qu’il existe une corrélation positive entre la taille des individus et la quantité de ressource disponible. Les corps de grandes tailles facilitent l’accumulation de réserves et ont un métabolisme plus lent. Ces caractéristiques leur confèrent une meilleure résistance à la faim.
Exemple de mammifères Nord-Américains - région Néarctique[8]:
La région Néarctique est une écozone s'étendant du Nord-Américain au Nord du Mexique, et comprenant le Groenland.
La biomasse des individus est directement liée à leur réserve en graisse. Le métabolisme des organismes les plus trapus est plus lent que celui des petits mammifères. Ainsi, les mammifères les plus gros, et donc au métabolisme le plus lent, ont une meilleure résistance à la famine que les plus petits. La disponibilité des ressources est donc également un facteur important de la variabilité de la taille. Il est très dur de séparer l'influence de la température et la disponibilité en ressources car ils sont liés au moins pour les végétaux.
Validité de la loi
Les données géographiques dont sont issues les études peuvent être biaisées. Par exemple, chez les Squamates, la majorité des données viennent de l’hémisphère Nord et de régions tempérées alors que les données sur les Chéloniens par exemple, viennent toutes de l’hémisphère Nord[8].
De plus, malgré une multitude d'études visant à démêler les mécanismes contrôlant la variation géographique de la taille corporelle, très peu tentent d'évaluer la validité de ces modèles de variation[7].
Il faut aussi prendre en compte que la majorité des études menées sur les questions de la règle de Bergmann utilisent des régressions ou des corrélations qui peuvent ne pas être linéaires[8].
L’article de Salewski[15] apporte des recommandations pour utiliser le terme « loi de Bergmann » car celui-ci peut porter à confusion avec les nombreuses études portant sur ce sujet :
- La loi de Bergmann inclut la thermorégulation, il ne faudrait donc pas prendre en compte d’autres facteurs.
- Les études portant sur les ectothermes ne devraient pas être liées à la loi de Bergmann.
- Les études sur la loi ne devraient se limiter à aucune unité taxonomique.
- Les études sur la validité de la loi devraient se concentrer sur le mécanisme plutôt que sur la recherche de tailles spécifiques.
Notes et références
- BLACKBURN, Tim M. et HAWKINS, Bradford A., 2004. Bergmann’s rule and the mammal fauna of northern North America. Ecography. [en ligne]. 2004. Vol. 27, p. 715-724. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/pdf/10.1111/j.0906-7590.2004.03999.x?casa_token=xUrVguV0h4wAAAAA:blv7N7whYGwnDkhTc0wtxTB9Hj4VGQmCoJcfAQ3VvsifY9I4FIe8Fnx3DXQR8xV_Ug0M-YJc2eW8
- BLACKBURN, Tim M., GASTON, Kevin J. et LODER, Natasha, 1999. Geographic gradients in body size: a clarification of Bergmann’s rule. Diversity and Distributions. [en ligne]. 1999. Vol. 5, p. 165‑174. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1046/j.1472-4642.1999.00046.x
- MEIRI, Shai, 2011. Bergmann’s Rule - what’s in a name? Global Ecology and Biogeography. [en ligne]. 2011. Vol. 20, p. 203‑207. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1111/j.1466-8238.2010.00577.x
- ROMANO, Andrea, SÉCHAUD, Robin et ROULIN, Alexandre, 2020. Generalized evidence for Bergmann’s rule: body size variation in a cosmopolitan owl genus. Journal of Biogeography. [en ligne]. 21 octobre 2020. Vol. 48, no. 1, p. 51‑63. DOI 10.1111jbi.13981. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/10.1111/jbi.13981
- SHELOMI, Matan et ZEUSS, Dirk, 2017. Bergmann’s and Allen’s Rules in Native European and Mediterranean Phasmatodea. Frontiers in Ecology and Evolution. [en ligne]. 5 avril 2017. Vol. 5, no. 25. DOI 10.3389/fevo.2017.00025. Disponible à l’adresse: https://www.frontiersin.org/articles/10.3389/fevo.2017.00025
- BROMMER, J. E., HANSKI, I. K., KEKKONEN, J. et VÄISÄNEN, R. A., 2014. Size differentiation in Finnish house sparrows follows Bergmann’s rule with evidence of local adaptation. Journal of Evolutionary Biology. [en ligne]. 27 février 2014. Vol. 27, p. 737‑747. DOI 10.1111/jeb.12342. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1111/jeb.12342
- MEIRI, Shai et DAYAN, Tamar, 2003. On the validity of Bergmann’s rule. Journal of Biogeography. [en ligne]. 20 mars 2003. Vol. 30, p. 331‑351. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1046/j.1365-2699.2003.00837.x
- ASHTON, Kyle G. et FELDMAN, Chris R., 2007. Bergmann’s rule in nonavian reptiles: turtles follow it, lizards and snakes reverse it. Evolution. [en ligne]. 9 mai 2007. Vol. 57, no. 5, p. 1151‑1163. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.0014-3820.2003.tb00324.x
- OISHI, Takuya, URAGUCHI, Kohji, ABRAMOV, Alexei V. et MASUDA, Ryuichi, 2010. Geographical Variations of the Skull in the Red Fox Vulpes vulpes on the Japanese Islands: An Exception to Bergmann’s Rule. Zoological Science. [en ligne]. 1er décembre 2010. Vol. 27, no. 12, p. 939‑945. Disponible à l’adresse: https://bioone.org/journals/zoological-science/volume-27/issue-12/zsj.27.939/Geographical-Variations-of-the-Skull-in-the-Red-Fox-Vulpes/10.2108/zsj.27.939.full
- MARTINEZ, Pablo A., MARTI, Dardo A., MOLINA, Wagner F. et BIDAU, Claudio J., 2013. Bergmann’s rule across the equator: a case study in Cerdocyon thous (Canidae). Journal of Animal Ecology. [en ligne]. 2013. Vol. 82, p. 997‑1008. DOI 10.1111/1365-2656.12076. Disponible à l’adresse: https://besjournals.onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2656.12076
- JOHNSON, David S., CROWLEY, Cynthia, LONGMIRE, Katherine, NELSON, James, WILLIAMS, Bethany et WITTYNGHAM, Serina, 2019. The fiddler crab, Minuca pugnax, follows Bergmann’s rule. Ecology and Evolution. [en ligne]. 2019. Vol. 9, p. 14489‑14497. DOI 10.1002/ece3.5883. Disponible à l’adresse: https://www.jstor.org/stable/pdf/10.1086/660834.pdf
- Atkinson, David 1994. Temperature and Organism Size - A Biological Law of Ectotherms?. Advances in Ecological Research, vol. 25, 1994, p. 1-58 DOI 10.1016/S0065-2504(08)60212-3, lire en ligne [sur researchgate.net]).
- GOHLI, Jostein et VOJE, Kjetil L., 2016. An interspecific assessment of Bergmann’s rule in 22 mammalian families. BMC Evolutionary Biology. [en ligne]. 19 octobre 2016. No. 16, p. 222‑234. DOI 10.1186/s12862-016-0778-x. Disponible à l’adresse: https://bmcecolevol.biomedcentral.com/articles/10.1186/s12862-016-0778-x
- ADAMS, Dean C. et CHURCH, James O., 2007. Amphibians do not follow Bergmann’s rule. Evolution. [en ligne]. 17 novembre 2007. Vol. 62, no. 2, p. 413‑420. DOI 10.1111/j.1558-5646.2007.00297.x. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1111/j.1558-5646.2007.00297.x
- SALEWSKI, Volker et WATT, Cortney, 2017. Bergmann’s rule: a biophysiological rule examined in birds. Oikos. [en ligne]. 2017. Vol. 126, no. 2, p. 161‑172. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1111/oik.03698
Voir aussi
Il y aurait un lien entre la règle de Bergmann et d’autres règles biologiques comme la règle de Cope (l’augmentation de la taille corporelle au cours des générations) et la règle de Rapoport. Cela suggère que la règle de Cope chez les mammifères peut-être une composante de la règle de Bergmann, au moins pendant certaines périodes.
Une variante de la loi de Bergmann est la loi d’Allen. Celle-ci stipule que la taille relative des extrémités du corps des endothermes (membres, queues, oreilles, etc.) devrait être plus petite dans des environnements plus froids afin de réduire les coûts de la thermorégulation. Cette loi est surtout applicable pour les mammifères, les ectothermex et les oiseaux de manière intraspécifique.
Articles connexes
Bibliographie
- ADAMS, Dean C. et CHURCH, James O., 2007. Amphibians do not follow Bergmann’s rule. Evolution. [en ligne]. 17 novembre 2007. Vol. 62, no. 2, p. 413‑420. DOI 10.1111/j.1558-5646.2007.00297.x. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1111/j.1558-5646.2007.00297.x
- ASHTON, Kyle G. et FELDMAN, Chris R., 2007. Bergmann’s rule in nonavian reptiles: turtles follow it, lizards and snakes reverse it. Evolution. [en ligne]. 9 mai 2007. Vol. 57, no. 5, p. 1151‑1163. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.0014-3820.2003.tb00324.x
- [Bergmann 1847] Carl Bergmann, « Über die Verhältnisse der Wärmeökonomie der Thiere zu ihrer Grösse », Göttinger Studien, vol. 3, no 1, 1847, p. 595–708.
- BLACKBURN, Tim M., GASTON, Kevin J. et LODER, Natasha, 1999. Geographic gradients in body size: a clarification of Bergmann’s rule. Diversity and Distributions. [en ligne]. 1999. Vol. 5, p. 165‑174. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1046/j.1472-4642.1999.00046.x
- BLACKBURN, Tim M. et HAWKINS, Bradford A., 2004. Bergmann’s rule and the mammal fauna of northern North America. Ecography. [en ligne]. 2004. Vol. 27, p. 715‑724. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/pdf/10.1111/j.0906-7590.2004.03999.x?casa_token=xUrVguV0h4wAAAAA:blv7N7whYGwnDkhTc0wtxTB9Hj4VGQmCoJcfAQ3VvsifY9I4FIe8Fnx3DXQR8xV_Ug0M-YJc2eW8
- BROMMER, J. E., HANSKI, I. K., KEKKONEN, J. et VÄISÄNEN, R. A., 2014. Size differentiation in Finnish house sparrows follows Bergmann’s rule with evidence of local adaptation. Journal of Evolutionary Biology. [en ligne]. 27 février 2014. Vol. 27, p. 737‑747. DOI 10.1111/jeb.12342. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1111/jeb.12342
- GOHLI, Jostein et VOJE, Kjetil L., 2016. An interspecific assessment of Bergmann’s rule in 22 mammalian families. BMC Evolutionary Biology. [en ligne]. 19 octobre 2016. No. 16, p. 222‑234. DOI 10.1186/s12862-016-0778-x. Disponible à l’adresse: https://bmcecolevol.biomedcentral.com/articles/10.1186/s12862-016-0778-x
- HUSBY, Arild, HILLE, Sabine M. et VISSER, Marcel E., 2011. Testing Mechanisms of Bergmann’s Rule: Phenotypic Decline but No Genetic Change in Body Size in Three Passerine Bird Populations. The American Naturalist. [en ligne]. août 2011. Vol. 178, no. 2. DOI 10.1086/660834.
- JOHNSON, David S., CROWLEY, Cynthia, LONGMIRE, Katherine, NELSON, James, WILLIAMS, Bethany et WITTYNGHAM, Serina, 2019. The fiddler crab, Minuca pugnax, follows Bergmann’s rule. Ecology and Evolution. [en ligne]. 2019. Vol. 9, p. 14489‑14497. DOI 10.1002/ece3.5883. Disponible à l’adresse: https://www.jstor.org/stable/pdf/10.1086/660834.pdf
- MARTINEZ, Pablo A., MARTI, Dardo A., MOLINA, Wagner F. et BIDAU, Claudio J., 2013. Bergmann’s rule across the equator: a case study in Cerdocyon thous (Canidae). Journal of Animal Ecology. [en ligne]. 2013. Vol. 82, p. 997‑1008. DOI 10.1111/1365-2656.12076. Disponible à l’adresse: https://besjournals.onlinelibrary.wiley.com/doi/epdf/10.1111/1365-2656.12076
- MEIRI, Shai, 2011. Bergmann’s Rule - what’s in a name? Global Ecology and Biogeography. [en ligne]. 2011. Vol. 20, p. 203‑207. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1111/j.1466-8238.2010.00577.x
- MEIRI, Shai et DAYAN, Tamar, 2003. On the validity of Bergmann’s rule. Journal of Biogeography. [en ligne]. 20 mars 2003. Vol. 30, p. 331‑351. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1046/j.1365-2699.2003.00837.x
- OISHI, Takuya, URAGUCHI, Kohji, ABRAMOV, Alexei V. et MASUDA, Ryuichi, 2010. Geographical Variations of the Skull in the Red Fox Vulpes vulpes on the Japanese Islands: An Exception to Bergmann’s Rule. Zoological Science. [en ligne]. 1er décembre 2010. Vol. 27, no. 12, p. 939‑945. Disponible à l’adresse: https://bioone.org/journals/zoological-science/volume-27/issue-12/zsj.27.939/Geographical-Variations-of-the-Skull-in-the-Red-Fox-Vulpes/10.2108/zsj.27.939.full
- [Roberts 1953] Derek Frank Roberts, « Body weight, race and climate », American Journal of Physical Anthropology, vol. 11, no 4, 1953, p. 533–558 (PMID 13124471, DOI 10.1002/ajpa.1330110404, résumé).
- [Roberts 1978] (en) Derek Frank Roberts, Climate and Human Variability, Menlo Park, CA, Cummings, 1978, 2e éd. (1re éd. 1973, Addison-Wesley Publishing Company), 38 p. (présentation en ligne).
- ROMANO, Andrea, SÉCHAUD, Robin et ROULIN, Alexandre, 2020. Generalized evidence for Bergmann’s rule: body size variation in a cosmopolitan owl genus. Journal of Biogeography. [en ligne]. 21 octobre 2020. Vol. 48, no. 1, p. 51‑63. DOI 10.1111jbi.13981. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/10.1111/jbi.13981
- [Ruff 1994] Christopher B. Ruff, « Morphological adaptation to climate in modern and fossil hominids », Yearbook of Physical Anthropology, vol. 37, 1994, p. 65–107 (DOI 10.1002/ajpa.1330370605, lire en ligne [sur onlinelibrary.wiley.com]).
- SALEWSKI, Volker et WATT, Cortney, 2017. Bergmann’s rule: a biophysiological rule examined in birds. Oikos. [en ligne]. 2017. Vol. 126, no. 2, p. 161‑172. Disponible à l’adresse: https://onlinelibrary.wiley.com/doi/epdf/10.1111/oik.03698
- [Schreider 1950] Eugène Schreider, « Geographical distribution of the body-weight/body-surface ratio », Nature, vol. 165, no 4190, 1950, p. 286 (PMID 15410342, DOI 10.1038/165286b0, Bibcode 1950Natur.165..286S).
- SHELOMI, Matan et ZEUSS, Dirk, 2017. Bergmann’s and Allen’s Rules in Native European and Mediterranean Phasmatodea. Frontiers in Ecology and Evolution. [en ligne]. 5 avril 2017. Vol. 5, no. 25. DOI 10.3389/fevo.2017.00025. Disponible à l’adresse: https://www.frontiersin.org/articles/10.3389/fevo.2017.00025