Mémoire (psychologie)
En psychologie, la mémoire est la faculté de l'esprit d'enregistrer, conserver et rappeler les expériences passées. Son investigation est réalisée par différentes disciplines : psychologie cognitive, neuropsychologie, et psychanalyse.
Psychologie cognitive

Le courant cognitiviste classique regroupe habituellement sous le terme de mémoire les processus d'encodage, de stockage et de récupération des représentations mentales. Beaucoup de recherches sur la mémoire en psychologie cognitive consistent à repérer et à décrire ses différents composants. Pour ce faire, les psychologues se basent sur des données expérimentales et sur les symptômes manifestés par des patients cérébrolésés (données neuropsychologiques).
Modèle modal de la mémoire
Le plus influent des modèles structuraux de la mémoire est le modèle modal, qui divise la mémoire en trois sous-systèmes : registre sensoriel, mémoire à court terme et mémoire à long terme[1]. Ce modèle est une synthèse de nombreux résultats expérimentaux et représente la conception dominante de la mémoire humaine dans la psychologie cognitive de la fin des années 1960. Une formulation classique de ce modèle a été proposée par Atkinson et Shiffrin[2] (1968).
Les trois composantes de la mémoire dans le modèle modal sont :
- Le registre sensoriel : il peut retenir une grande quantité d'informations sous forme visuelle pendant un temps extrêmement court (quelques millisecondes). Ce processus est différent du phénomène de rémanence visuelle.
- La mémoire à court terme (MCT) (ou mémoire de travail) : elle contient un nombre limité d'éléments, stockés sous forme verbale pendant quelques secondes.
- La mémoire à long terme (MLT) correspond à notre conception intuitive de la mémoire. La MLT ne connait pas en pratique de limites de capacité ou de durée de mémorisation.
Pour Atkinson et Shiffrin, la probabilité de mémorisation en mémoire à long terme dépend uniquement de la durée de présence en mémoire à court terme.
La notion de mémoire à court terme a ensuite été profondément renouvelée par le concept de mémoire de travail.
La dissociation entre MCT et MLT a conduit à l'élaboration de plusieurs modèles postulant que la mémoire est séquentielle[1]. Par exemple, il est difficile de réciter l'alphabet à l'envers lorsque celui-ci a été appris dans l'ordre. Il est difficile d'inverser l'ordre d'une séquence mémorisée comme telle[3] - [4].
Mémoire de travail (MDT)
Pour le modèle modal, la MCT joue un rôle particulier dans la cognition et particulièrement dans l'apprentissage de nouvelles informations. Les preuves expérimentales de ce fonctionnement sont cependant limitées.
Modèle de Baddeley
Devant les difficultés de ce modèle, et particulièrement pour rendre compte des propriétés dynamiques de la MCT, Alan Baddeley et ses collègues ont proposé un nouveau modèle de la mémoire de travail composé d'un administrateur central et trois sous-systèmes;
- L'administrateur central : mécanisme attentionnel de contrôle et de coordination des systèmes esclaves (boucle phonologique et calepin visuo-spatial). Il intègre les informations issues des deux sous-systèmes et les met en relation avec les connaissances conservées en mémoire à long terme. Pour cela, il dispose d'une zone tampon épisodique, qui lui permet de regrouper les informations, qu'elles soient issues des impressions sensibles ou de la mémoire à long terme.
Les trois sous-composants du modèle de Baddeley et Hitch sont :
- La boucle phonologique (BP) : elle est capable de retenir et de manipuler des informations sous forme verbale.
- Le calepin visuo-spatial (CVS) : il est chargé des informations codées sous forme visuelle.
- Le tampon épisodique (TE) : il permet aux informations contenues en MDT de passer en MLT et inversement. Les informations restent accessibles à la MDT qui doit encore les manipuler[5].
Modèle de Cowan
Un autre auteur, Cowan (1988), a développé sa propre théorie et son propre modèle de la mémoire de travail. Selon Cowan, la mémoire de travail ne représente que la partie activée de la MLT. Cowan, au contraire de Baddeley, se situe donc dans une vision unitaire de la MDT. Autrement dit, il n'y aurait pas spécifiquement de différence structurelle, mais seulement des différences fonctionnelles qui permettraient de rendre compte des différents « modules » ou fonctionnements de la MDT. Selon cet auteur, la partie la plus activée de la mémoire de travail correspond à ce qu'il nomme le focus attentionnel. En effet, l'attention portée sur certaines des informations activées serait dépendante du degré d'activation de ces dernières, soit par la perception, sous la forme de stimuli, soit sous la forme d'informations récupérées par les phénomènes d'amorçage. En d'autres termes, moins une information serait activée, moins elle aura de chance de faire partie d'une représentation explicite, verbale ou imagée.
Les différents types de mémoires décrits par Baddeley trouveraient leur explication dans la quantité de ressources ou d'énergie cognitive qu'il serait possible de solliciter par l'ensemble du système cognitif. Ainsi, cette quantité d'énergie plus ou moins limitée serait dirigée vers des « pôles d'attraction » correspondant aux zones les plus « centrales » par rapport à un contexte occurrent : situation vécue, thématique, raisonnement particulier, domaine de connaissances. La centralité d'une information, ou item, se mesure proportionnellement à sa familiarité (fréquence d'occurrence) dans un domaine, et par sa connexité, ou le nombre et la force des relations qu'entretient l'item considéré avec les autres informations du même domaine.
La mémoire de Cowan est à proprement parler un modèle de type connexionniste et automatiste : il n'existe qu'une seule structure composée d'unités fortement inter-reliées entre elles couplées à une fonction énergétique, représentant l'activation, qui se situe dans certaines zones du réseau d'unité en fonction des besoins. Ce modèle est automatiste puisqu'il ne fait pas non plus appel à certaines structures de contrôle ou de supervision : les propriétés physiques et mathématiques du réseau, des unités et de la fonction énergétique suffisent à rendre compte de l'ensemble des éléments décrits par Baddeley.
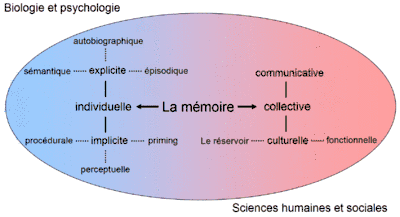
Mémoire à long terme : mémoire implicite et mémoire explicite
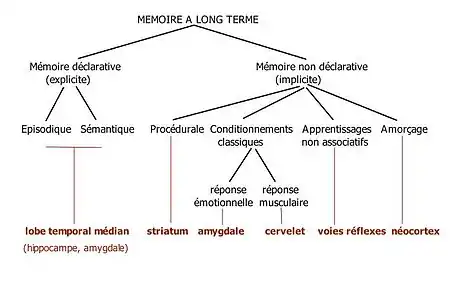
La mémoire à long terme est le sous-système mnésique ayant une capacité indéfinie de stockage dans le temps et en volume.
Les psychologues subdivisent la mémoire à long terme en deux sous-systèmes fonctionnels : la mémoire explicite (ou déclarative) et la mémoire implicite (ou non-déclarative). La distinction entre mémoire implicite et mémoire explicite concerne l'utilisation de la conscience lors du rappel. Les informations en mémoire explicite sont celles qui sont récupérées lors d'un rappel conscient, tandis que la mémoire implicite est celle des comportements et actions qui ne font pas intervenir la conscience.
Cette distinction entre mémoire implicite et explicite recouvre partiellement la distinction entre mémoire déclarative et non-déclarative. La mémoire déclarative est responsable de la mémorisation de toutes les informations sous forme verbale, c'est-à-dire celles qui peuvent être exprimées avec le langage. La notion de mémoire implicite et explicite généralise cette distinction à l'ensemble des natures de traitements d'information liés à la cognition humaine. Autrement dit, il existe des automatismes pour les informations verbales, imagées, sensitives et gestuelles autant qu'il existe des représentations mentales manipulables par la conscience et l'attention, sur lesquelles peuvent porter des décisions.
Une décision se réfère à la conscience : prendre une décision correspond à autoriser ou au contraire à inhiber un processus automatique préexistant. Au contraire des présupposés courants, la prise de décision ne « crée » pas à proprement parler de nouvelles informations, elle ne permet pas non plus d'en récupérer : elle permet simplement de porter un dernier processus de vérification sur des processus déjà déclenchés et des informations déjà activées et pré-structurées.
Comme pour l'ensemble des domaines liés à la cognition humaine, deux théories s'affrontent pour rendre compte de la distinction entre implicite et explicite : l'une structurelle et l'autre fonctionnelle. La théorie structurelle explique la différence implicite/explicite par une différence de nature physique : explicite et implicite correspondent à la sollicitation de modules et de structures cérébrales différentes. La théorie fonctionnelle suppose au contraire qu'il n'existe qu'un « tout » correspondant au support de la mémoire, mais aussi que ce tout est apte à différentes fonctions et au traitement de différentes natures d'information. Ce serait donc dans ce cas la sollicitation spécifique de différents contextes, fonctions et informations qui permettraient de rendre compte de la différence implicite/explicite.
Il existerait différentes formes de mémoire implicite et explicites.
Selon Squire[6], la mémoire implicite contiendrait les sous-formes de mémoires suivantes :
- la mémoire procédurale, qui permet l'acquisition et l'utilisation de compétences motrices comme faire du vélo ou pratiquer un sport ;
- la mémoire perceptive (dont une des manifestations serait l’amorçage), responsable de l'apprentissage des formes visuelles, des sons courants, etc. ;
- la mémoire des conditionnements classiques ;
- la mémoire des apprentissages non-associatifs, comme l'habituation et la sensibilisation.
La mémoire explicite, quant à elle, serait divisée en plusieurs sous-mémoires :
- la mémoire sémantique ;
- la mémoire épisodique.
Mémoire explicite : mémoire épisodique et mémoire sémantique
En ce qui concerne la mémoire à long terme explicite, plusieurs distinctions ont été établies entre la mémoire épisodique et la mémoire sémantique et entre la mémoire implicite (procédurale) et la mémoire explicite (déclarative). En outre, de très nombreuses recherches en psychologie cognitive portent sur les formes de représentations mentales utilisées en mémoire à long terme.
L'idée de la nécessité d'une mémoire sémantique contenant des connaissances générales pour la perception et la compréhension du langage a été suggérée par les recherches en intelligence artificielle. En psychologie, Endel Tulving a proposé en 1972, la distinction entre mémoire sémantique et mémoire épisodique (mémoire des événements de la vie personnelle), notamment pour rendre compte des symptômes de certains patients cérébrolésés présentant des troubles spécifiques à l'un de ces deux types de mémoire.
Modèle SPI de Tulving
Endel Tulving (1995) a proposé un modèle structural de la mémoire dans lequel il distingue cinq systèmes de mémoire organisés de façon hiérarchique, à la fois en termes d'origine phylogénétique et en termes de prépondérance au sein du système cognitif. On peut rappeler que Sherry et Schacter (1987) ont défini le terme de système de mémoire comme l'« interaction entre mécanismes d'acquisition, de rétention et de récupération, caractérisés par certaines règles opératoires (...), 2 systèmes (ou plus) se caractérisant par des règles fondamentalement différentes ».
Du plus ancien au plus récent, il considère les systèmes suivants, chacun d'eux nécessitant l'intégrité des systèmes précédents pour fonctionner :
- La mémoire procédurale : elle constitue selon ce modèle le plus ancien et le plus important système de mémoire ; son intégrité est nécessaire au fonctionnement des suivants
- Le système de représentation perceptive (SRP) : il contiendrait des ébauches perceptives des éléments constitutifs de la mémoire sémantique.
Ces deux premiers systèmes sont dits anoétiques puisqu'ils n'impliqueraient pas de prise de conscience de l'« objet ».
- La mémoire sémantique se réfère à l'ensemble des représentations des connaissances générales sur le monde.
- La mémoire primaire correspond à la MCT ou à la MDT. Ce système permet le maintien temporaire et la manipulation de l'information.
Ces deux systèmes sont dits noétiques puisqu'ils impliquent une prise de conscience des objets qu'ils traitent
- La mémoire épisodique concerne les représentations des événements situés dans le temps et dans l'espace (contexte). Ce système est dit auto-noétique parce qu'il implique une prise de conscience de l'objet et du sujet propre en tant qu'il perçoit l'objet.
Le modèle SPI (pour sériel, parallèle et indépendant) soutient que :
- L'encodage se fait de façon sérielle, dans un système après l'autre, item après item.
- Le stockage est parallèle, un élément pouvant être stocké dans plusieurs systèmes en même temps.
- La récupération se fait de manière indépendante, dans le système concerné.
Approche unitaire
Versace, Padovan et Nevers (2002) proposent une approche différente de la mémoire. Cette approche remet en question la conception en systèmes multiples de la mémoire, ainsi que la notion de représentation dans la valeur abstractive (l'objectif des systèmes sensoriels serait d'« abstraire » des invariants) que lui donne l'approche cognitiviste classique.
La conception en traces multiples considère que chaque confrontation à un événement entraine la création d'une trace mnésique, qui correspond strictement aux activations sensori-motrices (et notamment émotionnelles) provoquées par celui-ci. Ce serait l'accumulation de ces traces qui permettrait, à partir des confrontations répétées à un objet par exemple, dans une large étendue de contextes différents, d'extraire en quelque sorte un sens, recréé à chaque activation. Ce sens, qui n'est pas stocké en tant que tel, correspond en quelque sorte à l'ensemble des activations sensori-motrices liées à cet objet, en fonction du degré de liaison.
Mémoire et cognition située et distribuée
Alors que la plupart des modèles évoqués jusqu'à présent s'inscrivent dans la perspective du traitement de l'information en psychologie cognitive, certains auteurs proposent une vision radicalement différente de la cognition comme processus collectif inscrit dans l'environnement social et physique. Ces diverses perspectives sont généralement regroupées sous l'étiquette cognition située et distribuée.
Dans le domaine de la mémoire, on peut notamment citer le travail d'Edwin Hutchins (en) sur le pilotage d'avions de ligne et la navigation maritime. Il décrit, par exemple, comment le traitement (mémorisation, rappel, utilisation) d'un paramètre comme la vitesse de l'avion se distribue entre les deux membres de l'équipage et les outils à leur disposition dans le cockpit. Il suggère ainsi que les processus cognitifs ne sont pas des phénomènes purement individuels mais le résultat de l'activité coordonnée des participants et de leurs instruments.
Méta-mémoire
Pour expliquer la production du concept de l'écoulement temporel, il est nécessaire de faire appel au concept de méta-mémoire, ou méta-mnèse, c’est-à-dire une mémoire de la mémoire, caractérisée par le souvenir des variations de celle-ci. La méta-mnèse permet à l'esprit de s'abstraire du présent et d'imaginer un cours du temps en considérant la succession des souvenirs de ses états de mémoire ou plus précisément encore le souvenir des variations de sa mémoire. Cette propriété serait aussi nécessaire à la construction de la conscience de soi.
Oubli (Courbe théorique de l'oubli)
La « courbe théorique de l'oubli » suit une asymptote, ce qui signifie qu'un individu perd constamment de l'information (décroissance exponentielle) (bien qu'il n'oublie jamais tout totalement).
La psychologie sociale a montré que la communication joue cependant un rôle en renforçant les souvenirs dont on parle régulièrement et sélectivement, au détriment des autres (« oubli induit »)[7]. Plus un orateur évoque le souvenir d'une personne ou d'un évènement, plus - à la fois l'orateur et l'auditeur - s'en souviendront. Un phénomène de synchronisation et convergence de la mémoire peut ainsi apparaitre entre deux personnes ou au sein d'un groupe ou réseau social, qui a pour corollaire une convergence de l'« oubli induit »[7]. Les groupes peuvent ainsi et aussi adopter des souvenirs erronés ou déformer la mémoire individuelle et façonner des souvenirs collectifs[7]. Il a été expérimentalement montré qu'« une fois qu'un réseau est d'accord sur ce qui s'est passé, la mémoire collective devient relativement résistante à l'information concurrente ». Le phénomène de convergence de la mémoire (et donc des oublis) renforce la cohésion du groupe (d'une famille par exemple) et pourrait avoir un rôle social, mais il peut créer un fossé entre des groupes et poser problème quand il s'agit de rechercher la vérité (lors d'une procédure judiciaire notamment)[7]. « La mémoire forme l'identité de groupe, qui à son tour forme la mémoire, dans un cercle potentiellement vicieux (...) deux groupes peuvent converger vers des versions incompatibles du passé. Ces versions peuvent être conservées pour la postérité dans les statues et les livres d'histoire »[7].
La mémoire collective et historique est sélective (ainsi « l'Histoire du XXe siècle se souvient facilement des deux guerres mondiales, mais pas de la pandémie de grippe espagnole de 1918-20 qui a probablement tué plus qu'une seule de ces deux guerres »[7]). Elle évolue aussi avec le temps, notamment car les événements marquent plus la mémoire durant l'adolescence et chez le jeune adulte (« pics de réminiscence ») ; quand une génération grandit, les événements qui ont marqué ses membres durant leur jeunesse annulent les événements qui ont marqué la génération précédente, opérant une mise à jour de la mémoire collective[7].
Mémoire et neuroscience
La plasticité neuronale est fondamentale pour expliquer les diverses formes de mémoire. La règle de Hebb expose l'un des mécanismes les plus importants à l'origine de la mémorisation. Les zones de convergence-divergence sont vraisemblablement les lieux d'enregistrement et de reproduction des souvenirs, et d'autres formes de mémoire à long-terme.
Impact de certains neurotoxiques sur la mémoire
Certains médicaments et métaux lourds toxiques (ex plomb ou thallium) causent des troubles de la mémoire, souvent lentement réversibles (si l'intoxication n'a pas eu lieu in utero ou durant les premières années de la vie)[8].
Impact du stress sur la mémoire
Le stress joue un rôle important sur la formation de la mémoire ainsi que sur l’apprentissage. Lors des événements stressants, le cerveau relâche des hormones et des neurotransmetteurs (ex. : des glucocorticoïdes et des catécholamines) qui affectent les processus d'encodage en mémoire dans l’hippocampe. La recherche sur le comportement des animaux démontre que le stress chronique produit des hormones d’adrénaline qui ont une influence sur la structure de l’hippocampe dans les cerveaux des rats[9], par des biais ou mécanismes encore non compris.
Une étude expérimentale menée par les psychologues cognitivistes allemands L. Schwabe et O. Wolf montre comment l’apprentissage dans un environnement tendu réduit le rappel chez les humains aussi[10]. Dans leur étude, 48 étudiants universitaires (hommes et femmes) en bonne santé ont été répartis de façon aléatoire ou bien dans un groupe qui subit un test de stress ou dans un groupe témoin. Ceux qui ont participé au test de stress (le SECPT ou Socially Evaluated Cold Pressor Test) ont eu leur main immergée dans de l’eau glaciale pendant trois minutes durant lesquelles ils étaient surveillés et filmés. On a présenté aux deux groupes 32 mots pour mémoriser. Le jour suivant, les deux groupes ont été testés pour voir combien de mots ils ont pu se rappeler (le rappel libre) et combien ils ont pu reconnaître d'une liste plus longue (le taux de reconnaissance). Les résultats ont montré qu'il y avait une dépréciation nette du taux de reconnaissance pour le groupe qui avait subi le test de stress ; ceux-ci se rappelaient 30 % de mots de moins que le groupe témoin. Les chercheurs suggèrent que le stress vécu lors de l'apprentissage distrait les gens en détourant leur attention lors du processus d'encodage en mémoire.
La mémoire peut néanmoins être améliorée quand un lien est établi entre le matériel d’apprentissage et le contexte, même quand l’apprentissage se fait dans des conditions pénibles. Les psychologues cognitivistes Schwabe et Wolf ont montré que dans les situations où la conservation mnésique est contrôlée dans un contexte semblable ou congruent à la tâche d’apprentissage initiale (c'est-à-dire, dans la même chambre), les troubles de mémoire et les effets néfastes que pose le stress sur l’apprentissage peuvent être atténués[11].
Soixante-douze étudiants universitaires masculins et féminins et tous en bonne santé, affectés de façon aléatoire au test de stress SECPT ou au groupe témoin, avaient été demandés de se rappeler l’emplacement de 15 paires de cartes-images – une version informatique du jeu de cartes « Concentration » ou « Mémoire. » La salle dans laquelle l’expérience a eu lieu était infusée de parfum de vanille, puisque l’odorat est un indice fort pour la mémoire. Le contrôle de conservation mnésique eut lieu le jour suivant, soit dans la même salle, une nouvelle fois avec la présence du parfum de vanille, ou dans une autre salle sans ce parfum. La performance mnésique des sujets qui avaient ressenti du stress pendant la tâche objet-emplacement s’était considérablement détériorée lorsqu’ils avaient été contrôlés dans une salle non-familière et sans le parfum de vanille (un contexte non congruent). Toutefois, la performance mnésique des sujets stressés n’avait aucun signe de détérioration lorsque ces derniers étaient contrôlés dans la salle initiale avec le parfum de vanille (un contexte congruent). Tous les participants de l’expérience, stressés et non stressés, avaient une performance plus rapide lorsque l’apprentissage et les contextes de remémoration étaient semblables et congruents[12].
Ce travail de recherche portant sur l’impact du stress sur la mémoire peut avoir des implications pratiques quant à l’éducation, les témoignages par témoin oculaire et la psychothérapie : les étudiants peuvent avoir une meilleure performance quand leurs contrôles se font dans leur salle de classe habituelle plutôt que dans une salle d’examen ; les témoins oculaires pourraient mieux se souvenir des détails sur le lieu de l’évènement plutôt que dans une salle d’audience ; et la condition des personnes souffrant de névrose post-traumatique peut s’améliorer quand elles sont aidées à situer leurs souvenirs d’un événement traumatique dans un contexte approprié.
Psychopathologie
La psychopathologie de la mémoire comprend l'étude des troubles mnésiques organiques (maladies diverses, démences, etc.) et ceux qui résultent d'une affection, comme la dépression. La neuropsychologie se charge de l'étude des premiers[13].
Troubles mnésiques organiques
La plupart des maladies neurodégénératives touchant le cerveau ont un impact sur la mémoire.
Certaines infections bactériennes (ex. : maladie de Lyme) ou virales (ex. : phléborirose transmise par une tique)[14] affectent la mémoire (mémoire de court terme dans ces deux cas)[14].
Notes et références
- Sébastien Montel, 11 grandes notions de neuropsychologie clinique, Dunod, , 208 p. (ISBN 978-2-10-075099-3, lire en ligne), p. 40-41
- « Mémoire/Le modèle modal d'Atkinson et Shiffrin — Wikilivres », sur fr.wikibooks.org (consulté le )
- Alexia Audevart et Magaly Alonzo, Apprendre demain : Quand intelligence artificielle et neurosciences révolutionnent l'apprentissage, Dunod, , 224 p. (ISBN 978-2-10-080387-3, lire en ligne)
- (en) Ray Kurzweil, How to Create a Mind : The Secret of Human Thought Revealed, Prelude Books, , 336 p. (ISBN 978-0-7156-4595-6, lire en ligne)
- http://nbu.bg/cogs/events/2002/materials/Markus/ep_bufer.pdf.
- Squire, Memory systems of the brain : a brief history and current perspective, Neur of Learn. & mem, 82, 2004.
- Spinney L (2017), « How Facebook, fake news and friends are warping your memory ; Research on collective recall takes on new importance in a post-fact world » ; Nature 543, 168–170 (9 mars 2017) doi:10.1038/543168a.
- (en) Christopher Thompson, J. Dent et P. Saxby, « Effects of Thallium Poisoning on Intellectual Function », British Journal of Psychiatry, vol. 153, no 3, , p. 396–399 (ISSN 0007-1250 et 1472-1465, DOI 10.1192/bjp.153.3.396, lire en ligne, consulté le )
- (en) Conrad, C.D., « A critical review of chronic stress effects on spatial learning and memory. », Neuro-Psychopharmacology & Biological Psychiatry, vol. 5, no 34, , p. 742-755 (DOI 10.1016/j.pnpbp.2009.11.003).
- (en) Schwabe, L. et Wolf, O.T., « Learning under stress impairs memory formation », Neurobiology of Learning and Memory, vol. 2, no 93, , p. 183-188 (DOI 10.1016/j.nlm.2009.09.009).
- (en) Schwabe, L. et Wolf, O.T., « The context counts: Congruent learning and testing environments prevent memory retrieval impairment following stress », Affective & Behavioral Neuroscience, vol. 3, no 9, , p. 229-236 (DOI 10.3758/CABN.9.3.229).
- (en) Schwabe, L., Bohringer, A. et Wolf, O.T., « Stress disrupts context-dependent memory », Learning and Memory, vol. 2, no 16, , p. 110-113 (DOI 10.1101/lm.1257509).
- (en) Roger Gil, Neuropsychologie, Masson, coll. « Abrégés », , 496 p. (ISBN 978-2-294-08870-4 et 2-294-08870-0).
- William L. Nicholson, Ph.D., chief, pathogen biology and disease ecology activity, National Center for Emerging and Zoonotic Infectious Diseases, U.S. Centers for Disease Control and Prevention; Bruce Hirsch, M.D., attending physician, infectious diseases, North Shore University Hospital, Manhasset, N.Y.; Gary P. Wormser, M.D., chief, infectious diseases, New York Medical College and Westchester Medical Center, Valhalla, N.Y.; A New Phlebovirus Associated with Severe Febrile Illness in Missouri ; New England Journal of Medicine ; Aug. 30, 2012, .
Voir aussi
Articles connexes
- Classement thématique des neurosciences
- HM (patient)
- Maladie d'Alzheimer
- Mémoire (psychanalyse)
- Mémoire (sciences humaines)
- Mémoire eidétique
- Mini Mental State
- Mnémotechnique
- Myofasciite à macrophages
- Neuropsychologie, Amnésie, Démence
- Syndrome des faux souvenirs
- Zone de convergence-divergence
- Alain Lieury chevalier de l'ordre des Palmes académiques ayant travaillé sur la mémoire
- Courbe de l'oubli de Hermann Ebbinghaus
- Courbe d'apprentissage
Bibliographie
- (en) Alan Baddeley, Essential of Human Memory, Psychology Press, Hove, .
- (en) Alan Baddeley, La mémoire humaine, Grenoble, Presses Universitaires de Grenoble, .
- R. Versace, B. Nevers & C. Padovan, La mémoire dans tous ses états, Marseille, Solal, .
- Israël Rosenfield, L'invention de la mémoire (trad. A-S Cismareso), Paris, Flammarion, Revue critique de l'ensemble des approches anciennes et plus récentes de la mémoire, des théories localisationnistes au darwinisme neuronal d'Edelmann, en passant par la psychanalyse et le connexionnisme. Très documenté, synthétique, accessible à tout lecteur sans prérequis, et très rigoureux sur le plan théorique.
- Alain Lieury, Psychologie de la mémoire : Histoire, théories, expériences, Dunod, Paris, .
- Georges Chapouthier, Biologie de la mémoire, Paris, Odile Jacob, .
- (en) Atkinson Richard & Shiffrin Richard, « Human memory: A proposed system and its control processes », dans K. Spence et J. Spence (dir.), The Psychology of Learning and Motivation: Advances in Research and Theory, vol. 2, New York, Academic Press, , p. 89-195.
- (en) Baddeley Alan, « Working memory : Looking back and looking forward », Nature reviews, vol. 4, , p. 829-839.
- (en) Sherry, D.F., & Schacter, D.L., « The evolution of multiple memory systems », Psychological Review, no 94, , p. 439-454.
- (en) Tulving Endel, « Organisation of memory: quo vadis? In M.S. Gazzaniga (Ed) », The cognitive neurosciences, Cambridge, Mass: MIT Press, , p. 839-847.
- (en) Conrad, C.D., « A critical review of chronic stress effects on spatial learning and memory », Neuro-Psychopharmacology & Biological Psychiatry, vol. 5, no 34, , p. 742-755 (DOI 10.1016/j.pnpbp.2009.11.003).
- (en) « Learning under stress impairs memory formation », Neurobiology of Learning and Memory, vol. 2, no 93, , p. 183-188 (DOI 10.1016/j.nlm.2009.09.009).
- (en) Schwabe, L., & Wolf, O.T., « The context counts: Congruent learning and testing environments prevent memory retrieval impairment following stress. », Affective & Behavioral Neuroscience, vol. 3, no 9, , p. 229-236 (DOI 10.3758/CABN.9.3.229).
- (en) Schwabe, L., Bohringer, A., & Wolf, O.T., « Stress disrupts context-dependent memory », Learning and Memory, vol. 2, no 16, , p. 110-113 (DOI 10.1101/lm.1257509).
- Mewtow, Mémoire,
 .
.
Liens externes
- Ressources relatives à la santé :
- (en) Medical Subject Headings
- (no + nn + nb) Store medisinske leksikon
- Ressource relative à la recherche :
- Ressource relative à la littérature :
- Notices dans des dictionnaires ou encyclopédies généralistes :