Lisowicia
Lisowicia bojani

Lisowicia est un genre éteint de très grands thérapsides dicynodontes, avant vécu durant le Trias supérieur, entre 210 et 205 millions d'années avant notre ère dans ce qui est aujourd'hui la Pologne, et dont une seule espèce est connue, Lisowicia bojani. Il s'agit d'un des plus grands synapsides non-mammaliens jamais identifié à ce jour, en rivalité avec le géant Cotylorhynchus : on estime qu'il devrait peser entre 5 et 6 tonnes pour une taille comparable à celle des éléphants d'Asie. C'est aussi l'un des dicynodontes les plus récents connus, vivant peu avant leur extinction à la fin du Trias. Les fossiles de l'animal sont connus depuis 2008, mais le genre n'est officiellement décrit pendant une autre décennie à la fin de 2018.
Lisowicia est unique parmi les dicynodontes pour sa posture droite avec les quatre membres tenues debout directement sous son corps. Ceci est similaire aux membres de certains mammifères actuels et des dinosaures, contrairement aux postures tentaculaires et semi-dressées typiques de tous les autres dicynodontes (et de tous les autres synapsides non-mammaliens), et partage de nombreuses caractéristiques de ses membres ayant évolué indépendamment avec les grands mammifères. Lisowicia est le plus grand animal de son environnement, et est comparable en taille et en écologie aux sauropodomorphes « prosauropodes », qui occupent par ailleurs le rôle de grands herbivores à haute navigation dans la plupart des écosystèmes du Trias supérieur. Avant la découverte de Lisowicia, il est suggéré que les dicynodontes auraient été surpassés par les sauropodomorphes et serait incapables d'atteindre des tailles corporelles aussi grandes. Lisowicia peut avoir développé sa grande taille en réponse à l'absence de sauropodomorphes dans son écosystème dans le sud de l'actuelle Pologne, ou bien peut avoir agi comme un concurrent direct pour eux.
Découverte
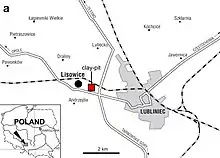
Les fossiles de Lisowicia ont été découverts dans la carrière d'argile de Lipie Śląskie à Lisowice, un village du sud de la Pologne près de la ville de Lubliniec, en Silésie. Les lits fossilifères de Lipie Śląskie se composent principalement de mudstones verts. L'âge de la carrière d'argile de Lipie Śląskie a été difficile à déterminer, avec différentes méthodes biostratigraphiques de datation fournissant des résultats variables. Le site a généralement été daté du Rhétien, ce qui ferait de Lisowicia l'un des dicynodontes plus récent au monde. Cependant, les paléontologues Grzegorz Racki et Spencer Lucas ont affirmé que le site était plus ancien et daté du stade Norien moyen sur la base de la biostratigraphie de grands vertébrés, dont Lisowicia. Ils ont plutôt suggéré que Lisowicia n'était pas significativement plus jeune que les autres dicynodontes et avait le même âge que les derniers genres connus ailleurs dans le monde[3].
Un échantillon de zircons détritiques de la fosse d'argile de Lipie Śląskie a été daté par radiométrie pour déterminer l'âge maximal de dépôt pour les lits de fossiles. Le plus jeune grain de zircon a enregistré un âge d'environ 211 ± 3 millions d'années à la fin du Norien, mais en raison de la différence entre la formation des cristaux de zircon et le dépôt sous forme de sédiment, il est probable que Lipie Śląskie soit plus jeune que cela, estimations du Norien au Rhétien ancien[4].
Les vestiges de Lisowicia ont été découverts pour la première fois en 2006 par les paléontologues Jerzy Dzik, Tomasz Sulej et Grzegorz Niedźwiedzki, qui les ont initialement confondus avec les os d'un dinosaure sauropodomorphe en raison de leur taille massive. Ils ont ensuite reconnu les fossiles comme ceux d'un dicynodonte géant en 2008, alors connus à partir d'un squelette partiellement articulé et de divers os isolés d'au moins deux individus. Entre 2007 et 2014 , plus de 1000 os de Lisowicia ont été collectés dans la carrière d'argile[5] - [6] - [7].
_Lisowicia_bojani.jpg.webp)
Lisowicia ne serait pas officiellement nommé et publié en tant que nouveau taxon avant janvier 2019, bien que le nom et la description aient été publiés en ligne en novembre 2018, avant la date de publication officielle. Le nom de genre fait référence au nom du village Lisowice où les spécimens originaux ont été trouvés, alors que le nom spécifique bojani mentionne Ludwig Heinrich Bojanus (1776-1827), un médecin et naturaliste allemand qui a fait un travail de pionnier dans l'anatomie comparée et la paléontologie[5]. Les fossiles connus de Lisowicia sont conservés dans les collections de l'Institut de paléobiologie de l'Académie polonaise des sciences et du Département de paléobiologie et d'évolution de l'Université de Varsovie, en Pologne.
D'autres restes d'un dicynodonte polonais géant ont été découverts près de la ville de Woźniki en 2007 et ont été signalés plus tard en 2010[8]. Ces restes avaient été considérés comme appartenant au même taxon que le dicynodonte de Lipie Śląskie, bien qu'ils n'aient pas été explicitement mentionnés comme appartennant à Lisowicia lorsqu'il a été décrit en 2019, et leur affinité n'est donc pas claire[9].
Description
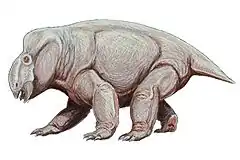
Bien que très grand pour un dicynodonte, Lisowicia ressemble encore beaucoup à d'autres stahleckeriidés du Trias, comme son proche parent Placerias d'Amérique du Nord. C'était un quadrupède fortement bâti avec un corps rond en forme de tonneau, une grosse tête avec des mâchoires à bec et un cou court, des membres trapus et une queue très courte[9].
Le crâne est incomplet et n'est connu que par diverses pièces individuelles isolées, mais elles suffisent à déterminer qu'il avait un crâne similaire à celui d'autres stahleckeriidés. La fenêtre temporale était large, et l'arrière du crâne était dessiné dans une haute crête entre eux, fournissant une attache étendue des muscles de la mâchoire à l'arrière du crâne. Le museau était relativement allongé comme chez les autres stahleckeriidés, et avait un bec édenté à l'avant. Comme certains autres dicynodontes du Trias, Lisowicia était complètement édenté et n'avait même pas les défenses typiques de la plupart des dicynodontes. Au lieu de cela, il avait une paire de projections triangulaires courtes et épaisses du maxillaire de la mâchoire derrière le bec appelé processus caniniformes, similaires à ceux de l'Ischigualastia et d'autres stahleckeriidés apparentés. De même, la comparaison des proportions de l'os carré à l'arrière du crâne à celles d'Ischigualastia suggère que Lisowicia avait un crâne large d'environ 43 centimètres de large entre chaque carré au niveau des articulations de la mâchoire[6].
Sa colonne vertébrale et ses côtes sont typiques des dicynodontes, cependant, les vertèbres cervicales du cou sont remarquables pour avoir des creux inhabituels creusés dans leurs côtés. Ceci est similaire aux trous dans certaines vertèbres de dinosaures qui sont associés à un système de sacs aériens et à un système respiratoire semblable à celui d'un oiseau. Cependant, contrairement aux dinosaures, Lisowicia manque d'ouvertures dans ses vertèbres pour les sacs aériens et l'intérieur de la vertèbre est solide. Ce trait est unique à Lisowicia parmi les dicynodontes, et sa fonction n'est pas claire. Les os du bassinet le membre postérieur sont très robustes par rapport aux autres dicynodontes en raison de la taille massive de l'animal (par exemple, le plus grand fémur connu mesure 80 centimètres de long), mais sont par ailleurs similaires à ceux des autres stahleckeriidés. Comme les autres stahleckeriidés, les pattes postérieures étaient également maintenues droites sous le corps[9].
Membre anterieurs
Les membres antérieurs de Lisowicia sont très distinctifs parmi les dicynodontes. En effet, leur construction est unique non seulement au groupe, mais aux synapsides non mammaliens dans leur ensemble et partage des caractéristiques avec les membres des mammifères et des dinosaures. Les membres antérieurs de Lisowicia sont tenus complètement dressés sous le corps, avec une articulation du coude qui est dirigée vers l'arrière et ne permet donc que l'avant-bras de se balancer d'avant en arrière dans une démarche parasagittale. Cet arrangement se trouve dans les membres postérieurs de divers autres grands Kannemeyeriiformes du Trias, mais ils conservent tous des membres antérieurs tentaculaires avec des coudes arqués vers l'extérieur et des articulations qui permettent le radius et le cubitus de tourner à chaque pas dans une posture allongée.
Pour atteindre cette posture, les membres antérieurs de Lisowicia ont dû subir plusieurs changements anatomiques importants par rapport à la condition normale des dicynodontes. Sur l'humérus, l'articulation de l'épaule et le coude sont parallèles l'un à l'autre, contrairement à l'humérus en rotation des dicynodontes tentaculaires, de sorte que l'humérus est maintenu droit sous l'épaule et directement au-dessus du radius et du cubitus. L'avant-bras lui-même est également exceptionnellement court par rapport aux autres dicynodontes puisque le coude est maintenant positionné plus bas, directement sous le corps et plus près du sol. La musculature du membre antérieur a également été réorganisée pour faciliter sa posture et sa démarche droites, fonctionnant désormais pour tirer le membre vers l'avant et vers l'arrière et perdant la capacité de faire pivoter le haut du bras comme il le ferait dans une foulée tentaculaire[9].
Taille

Bien que le squelette soit incomplet, la taille massive de divers éléments de son squelette indique que Lisowicia était un très gros animal, plus gros que tout autre dicynodontes connue. En comparant ses os à d'autres dicynodontes, la longueur de son corps a été estimée à environ 4,5 mètres de long et avec une hauteur de hanche de plus de 2 mètres[10]. Son poids corporel a été initialement calculé à environ 9,33 tonnes à partir de la circonférence de son fémur et de son humérus, une approximation pour estimer le poids que les membres devaient supporter. Cependant, les tiges de ces os sont d'une robustesse et d'une épaisseur disproportionnées chez Lisowicia, même pour sa taille, et son poids corporel a donc probablement été surestimé[9] - [11].
Son poids corporel a ensuite été estimé de manière plus fiable en modélisant sa masse à partir du volume total estimé de son corps. Ces estimations variaient en fonction de la circonférence de sa cage thoracique et de la quantité de tissus mous modélisés autour du squelette, avec un poids moyen global de 5,87 tonnes, et une estimation la plus basse avec un minimum de graisse corporelle et d'autres tissus à 4,87 tonnes et un maximum de 7,02 tonnes à son plus volumineux. Ces volumes sont comparable à celles des Éléphants d'Afrique modernes, faisant confortablement de Lisowicia le plus grand thérapside non-mammalien connu à ce jour ainsi que le plus grand synapside datant d'avant l'apparition des grands mammifères à l'Éocène (comme les brontothères). Par comparaison, le grand dinocéphale Tapinocaninus a été estimé de la même manière à peser seulement 892 kilos en moyenne[11] - [12].
Classification
Lisowicia est un membre des Kannemeyeriiformes, le groupe auquel appartiennent la plupart des dicynodontes datant du Trias, et fait partie de la famille des Stahleckeriidae, la dernière lignée de dicynodontes du Trias supérieur. En particulier, les analyses phylogénétiques de Sulej & Niedźwiedzki (2019) utilisant deux ensembles de données distincts ont toutes deux révélé que Lisowicia était étroitement liée au genre nord-américain Placerias dans la sous-famille des Placeriinae, avec le marocain Moghreberia. Un cladogramme simplifié de l'une des analyses portant sur les relations des Stahleckeriidae est reproduit ci-dessous :
| Stahleckeriidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||

Gigantisme et évolution des dicynodontes

Avant la découverte de Lisowicia, l'évolution de la taille des corps géants chez les sauropodomorphes du Trias était considérée comme un trait exclusif des dinosaures, car d'autres grands herbivores, y compris les dicynodontes, n'auraient pas atteint des tailles similaires. La taille importante de Lisowicia démontre que le gigantisme a également évolué chez les dicynodontes, et l'analyse de leur taille corporelle au fur et à mesure de leur évolution au cours du Trias supérieur démontre que, comme les sauropodomorphes, les dicynodontes du Trias augmentaient également en taille, culminant comme Lisowicia. En fait, les dicynodontes étaient systématiquement plus gros que les sauropodomorphes pendant le stade Carnien, et tous deux ont atteint des tailles gigantesques similaires durant le Norien et le Rhétien avec Lisowicia. L'évolution du gigantisme chez les sauropodomorphes et les Lisowicia à des périodes similaires suggère que des facteurs écologiques ont peut-être permis et encouragé les deux lignées à atteindre des tailles géantes, par opposition aux traits spécifiques inhérents aux dinosaures sauropodomorphes comme cela avait été suggéré précédemment[9].
L'évolution du gigantisme chez les dicynodontes a été un processus graduel, s'étalant sur environ 20 millions d'années, et a acquis une restructuration majeure du système musculo-squelettique pour atteindre la taille et la posture de Lisowicia. Il est possible que le gigantisme chez Lisowicia ait été motivé par l'augmentation de la taille corporelle des prédateurs, ou comme une adaptation métabolique pour obtenir plus de nutrition de leur nourriture en augmentant la taille de leur intestins[13] - [14]. Les paléontologues souligne également que l'évolution des dicynodontes au Trias supérieur est encore mal comprise. Auparavant considérée comme géographiquement restreinte et en déclin, Lisowicia démontre que les dicynodontes étaient à la fois encore géographiquement répandus et continuaient à se diversifier dans le Trias supérieur, en particulier des traits évoluant pour soutenir des tailles corporelles massives[9].
Paléobiologie
Alimentation et régime
Lisowicia était un herbivore, comme la plupart des autres dicynodontes, utilisant son bec relativement long pour récolter et mâcher, bien que sa grande taille implique qu'il se nourrissait de niveaux de végétation plus élevés que ce qui était typique pour le groupe. Les coprolithes attribués à Lisowicia indiquent qu'il se nourrissait principalement de végétation molle et de conifères. Cependant, certains coprolithes contiennent de grandes portions de matériel ligneux, un trait observé dans certains autres coprolithes de dicynodontes. Cela a été suggéré pour représenter la variation saisonnière de son régime alimentaire, peut-être lorsque leur régime préféré de végétation molle était hors saison[13].
Prédateurs

Des marques de morsure ont été trouvées sur les os de Lisowicia qui correspondent aux dents de l'archosaure prédateur contemporain Smok (en), et des fragments d'os de dicynodontes ont également été identifiés dans ses coprolithes proposés, ce qui implique que Lisowicia était un composant majeur du régime alimentaire du prédateur. La majorité des marques de morsures se trouvent sur des spécimens juvéniles de Lisowicia, et cette prépondérance a été suggérée pour indiquer que les Lisowicia juvéniles étaient préférentiellement chassés par Smok et qu'ils représentent une prédation active et pas seulement des restes charognards[14] - [15].
Croissance
Des études histologiques des os des membres de Lisowicia ont montré des caractéristiques d'un taux de croissance rapide en tant que juvéniles, similaires à d'autres grands dicynodontes. Cependant, alors que le taux de croissance des autres dicynodontes ralentissait au fur et à mesure, les os étudiés de Lisowicia n'ont montré aucun signe de ralentissement de leur croissance. La possibilité que les os appartenaient à des juvéniles avec une taille corporelle adulte encore plus grande que celle estimée a été exclue en partie sur la base de leur taille déjà massive, mais aussi parce que les os étaient bien ossifiés et matures sur le plan du développement. Au lieu de cela, il est plus probable que les Lisowicia ont grandi très rapidement sans aucune interruption jusqu'à atteindre leur taille adulte, car il n'y avait pas de lignes claires d'arrêt de croissance (LAG) qui indiquent par ailleurs des périodes de croissance plus lente et réduite. Les LAG ont peut-être été vraiment absents, ou ont peut-être été effacés par un remodelage étendu de l'os à l'âge adulte, qui soutiendraient tous deux une croissance rapide et permanente de Lisowicia, similaire aux mammifères et à certains dinosaures[9] - [5].
Latrines
Des coprolithes attribués à Lisowicia ont été découverts en accumulations abondantes dans des zones qui auraient été autour de l'eau stagnante. Ces assemblages sont similaires aux découvertes au Brésil censées représenter des latrines communales (en) faites par le dicynodonte Dinodontosaurus, et il est possible que Lisowicia ait eu un comportement similaire. Les latrines communales sont documentées chez les mammifères grégaires modernes et soutiennent l'idée que les dicynodontes comme Lisowicia vivaient en troupeaux et suggèrent même qu'ils avaient des comportements sociaux complexes comme les grands mammifères modernes[13] - [16].
Paléoécologie
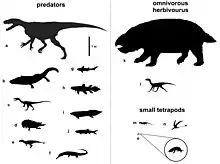
Lisowicia coexistait avec une grande variété d'animaux du Trias supérieur découvert à Lipie Śląskie. La majorité de ces animaux étaient des archosaures, dont des ptérosaures, des crocodylomorphes, le grand prédateur Smok ainsi que des dinosauromorphes dont un silesauridés[17]. D'autres petits reptiles comprennent un rhynchocéphale, un archosauromorphe indéterminé et un possible choristodère. Le seul autre thérapside connu pour coexister avec Lisowicia était Hallautherium, un petit mammaliaforme du sous-groupe éteint des morganucodontes, étroitement apparenté aux mammifères modernes et de plusieurs ordres de grandeur plus petit que Lisowicia. L'environnement d'eau douce humide abritait des amphibiens temnospondyles (un capitosaure et des plagiosauridés plus petits) ainsi qu'une abondance de poissons, notamment des cœlacanthes et des requins de la famille éteinte des hybodontidés[3] - [15].
L'environnement à Lipie Śląskie était humide et marécageux, comparable aux Everglades modernes[13], avec des rivières sinueuses et des lacs méandres lents, entourés d'une végétation abondante qui comprenait des conifères Cheirolepidiaceae, des fougères à graines telles que Lepidopteris et les cycadophytes Androstrobus et Beania. En revanche, les environnements voisins peuvent avoir été secs et arides, suggérés par les restes du conifère peut-être xérophyte Hirmeriella et des fragments de charbon de bois provenant d'incendies transportés par les rivières jusqu'à la localité[6] - [18] - [19].
L'absence de grands herbivores sauropodomorphes est curieuse, car ils sont présents dans de nombreux autres écosystèmes du Trias supérieur, y compris dans des localités voisines en Europe, comme Plateosaurus en Allemagne. Il est possible que des dicynodontes géants comme Lisowicia soient en concurrence directe avec les sauropodomorphes, mais il était également possible que leur absence n'ait aucun lien, et au lieu de cela, Lisowicia a atteint sa taille massive pour remplir le rôle écologique des grands herbivores brouteurs occupés par les sauropodomorphes dans d'autres écosystèmes[10].
Notes et références
Notes
Références
- BioLib, consulté le 16 octobre 2021
- Gigantic mammal “cousin” discovered
- G. Racki et S.G. Lucas, « Timing of dicynodont extinction in light of an unusual Late Triassic Polish fauna and Cuvier's approach to extinction », Historical Biology, , p. 1–11 (DOI 10.1080/08912963.2018.1499734)
- M. Kowal-Linka, E. Krzemińska et Z. Czupyt, « The youngest detrital zircons from the Upper Triassic Lipie Śląskie (Lisowice) continental deposits (Poland): Implications for the maximum depositional age of the Lisowice bone-bearing horizon », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 514, , p. 487–501 (DOI 10.1016/j.palaeo.2018.11.012)
- Gretchen Vogel, « A giant in the time of dinosaurs: Ancient mammal cousin looked like cross between a rhino and a turtle », Science, (DOI 10.1126/science.aaw1183, lire en ligne, consulté le )
- Jerzy Dzik, Tomasz Sulej et Grzegorz Niedźwiedzki, « A Dicynodont-Theropod Association in the Latest Triassic of Poland », Acta Palaeontologica Polonica, vol. 53, no 4, , p. 733–738 (DOI 10.4202/app.2008.0415)
- Nicholas St. Fleur, « An Elephant-Size Relative of Mammals That Grazed Alongside Dinosaurs », The New York Times, (lire en ligne, consulté le )
- T. Sulej, R. Bronowicz, M. Tałanda et G. Niedźwiedzki, « A new dicynodont–archosaur assemblage from the Late Triassic (Carnian) of Poland », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 101, no Special Issue 3—4, , p. 261–269 (DOI 10.1017/S1755691011020123)
- (en) Tomasz Sulej et Grzegorz Niedźwiedzki, « An elephant-sized Late Triassic synapsid with erect limbs », Science, vol. 363, no 6422, , p. 78–80 (ISSN 0036-8075, PMID 30467179, DOI 10.1126/science.aal4853)
- George Dvorsky, « During the Triassic, This Elephant-Sized Relative of Mammals Lived Alongside the Dinosaurs », Gizmodo, (lire en ligne, consulté le )
- Marco Romano et Fabio Manucci, « Resizing Lisowicia bojani: volumetric body mass estimate and 3D reconstruction of the giant Late Triassic dicynodont », Historical Biology, , p. 1–6 (ISSN 0891-2963, DOI 10.1080/08912963.2019.1631819)
- Marco Romano et Bruce Rubidge, « First 3D reconstruction and volumetric body mass estimate of the tapinocephalid dinocephalian Tapinocaninus pamelae (Synapsida: Therapsida) », Historical Biology, , p. 1–8 (ISSN 1029-2381, DOI 10.1080/08912963.2019.1640219)
- P. Bajdek, K. Owocki et G Niedźwiedzk, « Putative dicynodont coprolites from the Upper Triassic of Poland », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 411, , p. 1–17 (DOI 10.1016/j.palaeo.2014.06.013)
- G. Niedźwiedzki, P. Gorzelak et T. Sulej, « Bite traces on dicynodont bones and the early evolution of large terrestrial predators », Lethaia, vol. 44, no 1, , p. 87–92 (DOI 10.1111/j.1502-3931.2010.00227.x)
- Martin Qvarnström, Per E. Ahlberg et Grzegorz Niedźwiedzki, « Tyrannosaurid-like osteophagy by a Triassic archosaur », Scientific Reports, vol. 9, no 1, , p. 925 (PMID 30700743, PMCID 6353991, DOI 10.1038/s41598-018-37540-4)
- L.E. Fiorelli, M.D. Ezcurra, E.M. Hechenleitner, E Argañaraz, J.R.A. Taborda, M.J. Trotteyn, M.B. von Baczko et J.B. Desojo, « The oldest known communal latrines provide evidence of gregarism in Triassic megaherbivores », Scientific Reports, vol. 3, no 3348, (DOI 10.1038/srep03348)
- Edwin H. Colbert, « Variation in Coelophysis bauri », dans Dinosaur Systematics, Cambridge University Press, (lire en ligne), p. 81–90
- K. Jewuła, M. Matysik, M. Paszkowski et J. Szulc, « The late Triassic development of playa, gilgai floodplain, and fluvial environments from Upper Silesia, southern Poland », Sedimentary Geology, vol. 379, , p. 25–45 (ISSN 0037-0738, DOI 10.1016/j.sedgeo.2018.11.005)
- Z. Wawrzyniak, Morphology and Systematics of Fossil Vertebrates, DN Publisher, Wrocław, , 118—124 (OCLC 751467953, lire en ligne), « What could eat dicynodonts (Dicynodontia, Therapsida) from Lipie Śląskie? (Upper Silesia, Poland) – preliminary results of research of the Late Triassic macroflora »
Voir aussi
Articles connexes
Publication originale
Bibliographie
- (en) Jerzy Dzik, Tomasz Sulej et Grzegorz Niedźwiedzki, « A Dicynodont-Theropod Association in the Latest Triassic of Poland », Acta Palaeontologica Polonica, PAN et Institute of Paleobiology (d), vol. 53, no 4, , p. 733-738 (ISSN 0567-7920 et 1732-2421, OCLC 02051833, DOI 10.4202/APP.2008.0415, lire en ligne)
Liens externes
- (en) Référence BioLib : Lisowicia Sulej & Niedźwiedzki, 2018 † (consulté le )
- (en) Référence Paleobiology Database : Lisowicia Sulej & Niedźwiedzki, 2018 † (consulté le )
- Ressources relatives au vivant :