Astrocyte
Les astrocytes sont des cellules gliales du système nerveux central. Elles ont généralement une forme étoilée, d'où provient leur étymologie (origine grec) : Astro - étoile et cyte - cellule. Elles assurent une diversité de fonctions importantes, centrée sur le support et la protection des neurones. Ces cellules participent au maintien de la barrière hémato-encéphalique, régulent le flux sanguin, assurent l'approvisionnement en nutriments et le métabolisme énergétique du système nerveux, participent à la neurotransmission, à la détoxification du milieu extracellulaire notamment par capture du glutamate, et maintiennent la balance ionique du milieu extracellulaire. Les astrocytes jouent également un rôle dans la défense immunitaire, la réparation et la cicatrisation du cerveau ou de la moelle épinière après une lésion[2].

Les recherches récentes révèlent la complexité et l'importance de cette population cellulaire. Les astrocytes sont capables d'assurer une certaine forme de communication, reposant sur des vagues intracellulaires de Ca2+, et peuvent également libérer certains neurotransmetteurs (appelés gliotransmetteurs). Cette forme de communication, que l'on croyait spécifique aux neurones, leur confèrerait un rôle beaucoup plus actif dans le fonctionnement du cerveau, notamment sur la plasticité des communications neuronales. Il a également été montré que certaines sous-populations d'astrocytes ont des propriétés de cellules souches neurales et sont à la source du mécanisme de neurogenèse adulte.
Histoire des découvertes

Rudolf Virchow fut le premier, en 1858, à proposer le concept de « neuroglie » comme étant l'élément cellulaire interstitiel du système nerveux. Camillo Golgi observa par la suite les astrocytes et supporta également qu'ils constituaient la « glu » ou le support du cerveau. Le terme « astrocyte » fut utilisé pour la première fois en 1893, par Mihály Lenhossék, pour rendre compte de leur forme étoilée.
Les travaux de Kolliker et Andriezen distinguèrent par la suite les astrocytes fibreux et protoplasmiques. Néanmoins, ce ne fut qu'à partir de Ramón y Cajal et ses dessins très détaillés des composants du système nerveux, que les nombreuses formes et fonctions des astrocytes furent observées[2]. En effet, c'est en utilisant une coloration au chlorure d'or que Cajal marqua par hasard les filaments intermédiaires des astrocytes.
Ce n'est qu'en 1919 que les oligodendrocytes et les cellules microgliales furent séparées des astrocytes, distinguant ainsi les trois classes actuelles des cellules gliales du cerveau (une quatrième classe, les cellules NG2, semble émerger depuis les années 2000).
Désormais, de nombreuses études montrent la diversité des astrocytes et leurs rôles majeurs dans le développement, le fonctionnement et l'évolution du système nerveux.
Description
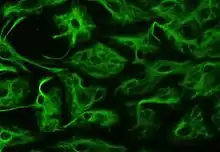
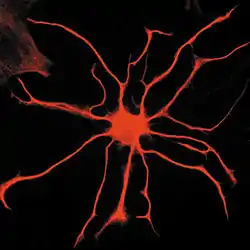
Il existe de nombreux types et formes d'astrocytes et il est très difficile de cerner la nature commune des astrocytes. Ces cellules ont tout d'abord été distinguées en fonction de leur morphologie :
- d'un côté, les astrocytes protoplasmiques de la substance grise présentant de nombreuses ramifications courtes ;
- de l'autre, les astrocytes fibreux de la substance blanche ayant moins de ramifications, mais celles-ci sont longues et lisses.
D'autres types d'astrocytes viennent s'ajouter à ces deux catégories :
- les astrocytes de la glie radiaire sont un type particulier de cellules gliales présent majoritairement durant le développement embryonnaire ;
- la glie de Bergmann au niveau du cervelet ;
- la glie de Muller au niveau de la rétine ;
- les cellules souches neurales adultes qui possèdent certaines propriétés d'astrocyte.
Les astrocytes ne possèdent pas de signature moléculaire spécifique et sont le plus souvent identifiés par leur morphologie ou leurs interactions avec les neurones ou les capillaires sanguins. Néanmoins la protéine GFAP (Glial Fibrilary Acidic Protein), un des composants de leurs filaments intermédiaires, est très largement utilisée comme marqueur des astrocytes, bien qu'elle ne soit pas exprimée dans tous les astrocytes et que d'autres types cellulaires l'expriment également[2].
Le corps cellulaire des astrocytes contient un noyau et un cytoplasme peu abondant. Les prolongements cytoplasmiques sont quant à eux très importants, parcourant notamment tout le neuropile. Ils forment aussi la glia limitans, film protecteur au contact des méninges, formé par les prolongements astrocytaires et isolant ainsi le système nerveux central.
En microscopie optique, il faut des colorations spéciales, métalliques à base d'argent pour pouvoir voir ces prolongements. Ils sont par ailleurs visualisés par immunohistochimie en utilisant des anticorps spécifiques, notamment celui détectant le GFAP. En microscopie électronique, on peut distinguer les organites classiques (mitochondrie, appareil de Golgi, reticulum endoplasmique). Les astrocytes ont également la particularité de présenter des grains de glycogène, forme sous laquelle ils stockent l'énergie apportée par les sucres.
Classification fonctionnelle
Les astrocytes de type I sont en contact avec les capillaires sanguins grâce à leurs pieds astrocytaires. Ils jouent un rôle actif dans le métabolisme et l'alimentation en lactate des neurones, via la glycolyse du glucose provenant des capillaires sanguins. La sécrétion de glutamate par le neurone favorise cette capture du glucose, ce qui permet de lier l'activité du neurone (le glutamate) à une demande accrue en énergie (le glucose).
Les astrocytes de type II, entourent le neurone et la fente synaptique. Le taux de couverture peut varier de 1 à 100 % suivant le type de neurone. On suppose qu'ils jouent un rôle dans la propagation du signal nerveux en agissant sur la dispersion des neurotransmetteurs. De plus, les astrocytes agissent sur la recapture des différents neurotransmetteurs, influant ainsi sur l'intensité d'un signal et sa durée. Le glutamate est, par exemple, recapturé par un cotransport avec le sodium[4].
Fonctions
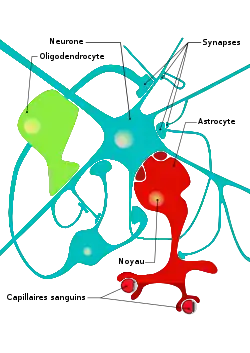
Les astrocytes assurent un grand nombre de fonctions vitales au fonctionnement et à la survie des neurones.
Structure tridimensionnelle
La première fonction supposée des astrocytes est celle de support physique. Leur densité dans le cerveau leur permettrait de constituer le réseau tridimensionnel qui assure le maintien des structures cérébrales et de leurs organisations. De plus, les astrocytes ne sont pas des cellules excitables comme les neurones (ils ne peuvent pas conduire de potentiels d'action) ce qui a dirigé les scientifiques à les considérer exclusivement comme des cellules support.
Rôle synaptique
Néanmoins, il s'est avéré que les astrocytes possèdent des propriétés bien plus importantes que le simple rôle de support. Au niveau de la synapse, les astrocytes interviennent dans le recyclage des neurotransmetteurs et produisent également certains nombres de coagonistes modulant l’efficacité de la synapse. Ils jouent donc un rôle actif dans la communication neuronale. Leur implication a amené à poser le « modèle de la synapse tripartite », composé d'un espace présynaptique, d'un espace post-synaptique et d'un espace astrocytaire. Notamment, la recapture du glutamate, un neurotransmetteur excitateur, par les astrocytes permet de réguler l'activité de la synapse et de prévenir l'excito-toxicité (à forte concentration, ou présent trop longtemps dans la fente synaptique, le glutamate devient toxique pour le neurone). Dans les astrocytes, le glutamate est transformé en glutamine et est ensuite recédé aux neurones qui les transforment en neurotransmetteurs (GABA). Les astrocytes produisent des transporteurs membranaires de glutamate, majoritairement GLAST et GLT-1, dont la proportion varie dans le système nerveux central et durant le développement. Les astrocytes ne présentent pas tous la même capacité à capturer le glutamate, il existe notamment une différence entre astrocytes fibreux et protoplasmique[2].
Vagues calciques
Les astrocytes ne sont pas excitables électriquement, mais ils sont excitables chimiquement. Sous certains stimuli — notamment en présence de certains neurotransmetteurs, comme le glutamate, l'ATP, le GABA, l'adénosine, la noradrénaline, l'acétylcholine, les prostaglandines ou les endocannabinoïdes — les astrocytes augmentent leur concentration intracellulaire de calcium (Ca2+). Cette augmentation se propage alors à travers les astrocytes voisins, en partie grâce à des jonctions communicantes (gap junctions), constituant une vague calcique. La genèse et les conséquences de cette augmentation calcique ne sont pas encore parfaitement connues, mais sont dépendantes du type et de la localisation de l'astrocyte et de l'activité de son environnement neuronal[2].
Modulation de l'activité
Les astrocytes réagissent à certains neurotransmetteur en augmentant leur concentration intracellulaire de calcium. En conséquence de cette augmentation, ils sont notamment capables de libérer différents gliotransmetteurs parmi lesquels le glutamate, l'ATP, l'adénosine, la D-sérine, le TNF-α et des eicosanoïdes, qui modulent à leur tour l'activité des cellules de la zone, notamment les autres astrocytes, les neurones, la microglie et les cellules vasculaires présentes. Un seul astrocyte peut être en contact avec plusieurs milliers de synapses d'une même zone. En libérant des gliotransmetteurs dans les fentes synaptiques, les astrocytes moduleraient l'activité électrique de plusieurs neurones et la plasticité de ces connexions. Néanmoins, l'action physiologique in situ des gliotransmetteurs n'a pas encore été prouvée et il pourrait également s'agir d'un biais lié à l'expérimentation[2].
Remodelage de circuits neuronaux
Des neuroscientifiques de l’école de médecine de l’université Stanford ont découvert un nouveau rôle joué par les astrocytes. Leurs conclusions ont été publiées dans la revue Nature du . Ces cellules redéfinissent les circuits en éliminant des contacts synaptiques un peu comme un sculpteur enlève le surplus de matière pour en faire une œuvre d’art. « Il s’agit d’une fonction entièrement nouvelle et inconnue encore des astrocytes, » a déclaré Ben Barres, professeur, titulaire de la chaire de neurobiologie et directeur de cette étude. L’auteur principal de cette étude, dans les équipes de recherche de Ben Barres, a été Won-Suk Chung. L’étude a été effectuée sur des tissus du cerveau de souris, mais cette découverte est susceptible de s’appliquer aussi à l’être humain selon Ben Barres… La découverte s'ajoute à d'autres analyses montrant qu'un cerveau adulte rénove, de façon constante, ses circuits cérébraux, et que les astrocytes sont des « maîtres-sculpteurs ». Les astrocytes jouent un rôle actif dans l'«élagage» des synapses. Les conclusions soulèvent également la question de savoir si des médicaments pourraient être conçus pour éviter la perte de cette capacité de remodelage, qui pourrait expliquer certaines maladies neurodégénératives telles que la maladie d’Alzheimer ou la maladie de Parkinson[5].
Fermeture de la période de plasticité cérébrale postnatale
Une étude de 2021, met en évidence le rôle crucial joué par les astrocytes dans la fermeture de la période de plasticité cérébrale essentielle pour le développement des facultés sensorielles et cognitives après la naissance[6]. L'étude s'est focalisée sur le cortex visuel de la souris[7].
Rôle vasculaire
En complément du prolongement synaptique, les astrocytes présentent un prolongement en contact avec la surface des capillaires qui irriguent le cerveau. Ces pieds vasculaires recouvrent complètement la surface des capillaires et participent à la barrière hémato-encéphalique, limitant les échanges entre le domaine vasculaire et le domaine neuronal. Bien que leur rôle dans le maintien de cette barrière ne soit pas prépondérant[8], il semble que l'activité des astrocytes contrôle le flux sanguin et qu'il y a donc une communication entre ces pieds astrocytaires et les cellules endothéliales des vaisseaux. Ce serait ainsi l'activité des astrocytes et non celle des neurones qui serait mesurée en IRM fonctionnelle[9].
C'est au travers de ces prolongements que les astrocytes captent également les nutriments essentiels aux fonctionnements du cerveau, premier consommateur énergétique du corps humain. Ils présentent notamment un mécanisme de glycolyse qui leur permet de fournir le lactate, le sucre nécessaire aux neurones.
Les astrocytes marginaux, dont les prolongements sont au contact des méninges et du liquide cérébrospinal, ont également un rôle de filtre.
Régulation de la neurogenèse
Les astrocytes participent à la régulation de la neurogenèse adulte, en sécrétant une série de facteurs (comprenant Wnt3, l'interleukine-1β, l'interleukin-6 et l'insulin-like growth factor binding protein 6) ainsi que par l'expression de certains facteurs membranaires. Certaines protéines, largement exprimées par les astrocytes, comme vimentin ou GFAP, influeraient sur les capacités de neurogenèse et de régénération du tissu nerveux, régulation impliquant la voie de signalisation Notch. Enfin, les astrocytes régulent également l'intégration et la survie des nouveaux neurones dans les circuits préexistants, en agissant notamment sur la création ou l'élimination de leurs synapses[10].
Astrogliose
Les astrocytes présentent plusieurs états d'activité. Lors d'une lésion ou d'une infection par exemple, ils sont activés et participent à la cicatrisation ou à la réaction immunitaire. C'est le mécanisme d'astrogliose.
Pathologies
La densité des astrocytes dans le système nerveux central et leur implication dans de nombreuses fonctions vitales font que ces cellules peuvent être impliquées dans de nombreux processus pathologiques.
Les changements s'opérant dans la structure et la physiologie des astrocytes sont souvent regroupés sous le terme d'activation ou réaction astrocytaire. C'est un ensemble de modifications du phénotype de ces cellules regroupant une modification de la synthèse et de la distribution de protéines du cytosquelette, la libération de molécules apparentées à celles impliquées dans l'inflammation ainsi qu'une perturbation de la capacité de ces cellules à réguler la composition extracellulaire : recapture de neurotransmetteurs, ions, etc. Cette activation est un processus physiologique de défense et de cicatrisation. Néanmoins, elle présente de nombreux défauts qui sont susceptibles de favoriser l'apparition ou l'aggravation de pathologies aiguës ou chroniques comme la sclérose en plaques. Notamment lors de l'astrogliose à la suite d'une lésion, les astrocytes comblent rapidement le vide laissé par la liaison, mais y forment une cicatrice qui empêche les axones de repousser et de rejoindre leur cible. Ce problème, souvent rencontré lors de paralysie à la suite d'une atteinte de la moelle épinière est un des axes majeurs de la recherche médicale en neurosciences. Celle-ci cherche comment forcer le système nerveux à créer une cicatrice perméable aux axones tout en maintenant la structure et les défenses actives. Les greffes de cellules gliales modifiées ou provenant d'autres parties du corps (cellules de Schwann, cellules engainantes olfactives, etc.) sont une des pistes actuelles prépondérantes.
Les astrocytes seraient également impliqués dans le développement de la maladie d'Alzheimer[11]. L'accumulation de β-amyloïde, caractéristique de cette pathologie dégénérative, perturbait l'excitabilité chimique des astrocytes, via une forte hausse de calcium intracellulaire au repos. Cette accumulation calcique semblerait être principalement due à une libération de calcium stocké dans le réticulum endoplasmique des astrocytes. Cela conduirait alors à une hyperexcitabilité de ces cellules nerveuses. Des vagues calciques plus importantes, ainsi que des oscillations à profil pathologique ont alors été observées chez la souris[12]. Outre cette augmentation de calcium intracellulaire, l'exposition à la β-amyloïde aurait également un impact sur l'expression de certaines protéines situées à la surface des astrocytes[13]. Par exemple, l'expression de mGluR5, un récepteur métabotropique des astrocytes, pourrait augmenter en présence de β-amyloïde. La stimulation de son expression a été détectée chez des astrocytes, à travers le niveau protéique ainsi que par l'ARNm spécifque, après avoir été mis en contact avec l'agrégat protéique[14]. Augmenter le nombre de récepteurs provoquerait alors une signalisation chimique d'une amplitude plus importante qu'en conditions physiologiques. Cette hausse de mGluR5 a été détectée dans le cortex cérébral de souris, chez des astrocytes réactivés associés à des plaques séniles[15].
Comme la majorité des types cellulaires, les astrocytes peuvent être à l'origine de tumeurs telles que l'astrocytome bénin, et l'astrocytome anaplasique qui se classent parmi les tumeurs cérébrales.
Origine
Chez l'humain, les astrocytes sont des dérivés de l'ectoblaste embryonnaire.
Notes et références
- Karin Pierre, Institut de Physiologie, UNIL, Lausanne: Alliance Européenne Dana pour le Cerveau (EDAB).
- NA Oberheim, SA Goldman and M Nedergaard, Heterogeneity of Astrocytic Form and Function ch.3 of Astrocytes: Methods and Protocols, Methods in Molecular Biology, vol. 814, Richard Milner (ed.) 2012.
- Alliance Européenne Dana pour le Cerveau (EDAB).
- MC Raff et al., Glial cell diversification in the rat optic nerve, Science 17 mars 1989, Vol. 243 no. 4897 pp. 1450-1455 DOI 10.1126/science.2648568.
- (en) Bruce Goldman, « Common brain cell plays key role in shaping neural circuits, study finds », sur le site med.stanford.edu.
- « Le rôle clé des astrocytes pour le développement cognitif », sur INSERM salle de presse, (consulté le ).
- DOI 10.1126/science.abf5273
- Kimelberg HK, Jalonen T, Walz W. Regulation of the brain microenvironment: transmitters and ions. In: Murphy S, editor. Astrocytes: pharmacology and function. San Diego, CA: Academic Press; 1993. p. 193–222. January 2008.
- (en) Swaminathan N, « Brain-scan mystery solved », Scientific American Mind, vol. Oct-Nov, , p. 7.
- Parpura et al. Glial cells in (patho)physiology, The Authors Journal of Neurochemistry, 2012.
- (en) Verkhratsky A, « Astroglial Calcium Signaling in Aging and Alzheimer's Disease », sur Cold Spring Harbor perspectives in biology, (PMID 31110130, consulté le )
- Kishore V. Kuchibhotla, Carli R. Lattarulo, Bradley T. Hyman et Brian J. Bacskai, « Synchronous hyperactivity and intercellular calcium waves in astrocytes in Alzheimer mice », Science, vol. 323, no 5918, , p. 1211–1215 (ISSN 0036-8075, PMID 19251629, DOI 10.1126/science.1169096, lire en ligne, consulté le )
- (en) Lim D et Ronco V, « Glial Calcium Signalling in Alzheimer's Disease », sur Reviews of physiology, biochemistry and pharmacology, (PMID 24935225, consulté le )
- (en) Christopher S. Casley, Viktor Lakics, Hyoung-gon Lee et Lisa M. Broad, « Up-regulation of astrocyte metabotropic glutamate receptor 5 by amyloid-β peptide », Brain Research, vol. 1260, , p. 65–75 (ISSN 0006-8993, DOI 10.1016/j.brainres.2008.12.082, lire en ligne, consulté le )
- (en) Andrea Wevers et Hannsjörg Schröder, « Nicotinic Acetylcholine Receptors in Alzheimer's Disease* », Journal of Alzheimer's Disease, vol. 1, nos 4-5, , p. 207–219 (ISSN 1387-2877, DOI 10.3233/JAD-1999-14-503, lire en ligne, consulté le )
Voir aussi
Bibliographie
- Science & vie, Nov-2005, « Nous avons un deuxième cerveau », 13 pages
- Cours de Biologie Cellulaire, de la faculté de médecine Joseph Fourier I de Grenoble
Articles connexes
Liens externes
- A Theory of Cortical Neuron-Astrocyte Interaction (en anglais)
- Astrocytes de rat
- La cellule nerveuse : Neurones et astrocytes
{kind=link}