Éclipse du darwinisme
Le terme d'éclipse du darwinisme, popularisé par Julian Huxley, désigne une phase de l'histoire de la biologie pendant laquelle le principe de l'évolution était largement admis, mais pas la sélection naturelle en tant que mécanisme explicatif. Des historiens des sciences, tel Peter Bowler, ont utilisé l'expression pour désigner une période s'étendant à peu près de 1880 à 1930, durant laquelle des mécanismes différents de ou complémentaires à la sélection naturelle sont étudiés en détail, de nombreux biologistes voyant dans la sélection naturelle une supposition hâtive de Charles Darwin (1809-1882) ou minimisant son importance. L'expression « interphase du darwinisme » a été proposée, le terme d'éclipse pouvant laisser penser, à tort, qu'une période d'intense recherche darwinienne l'avait précédée.

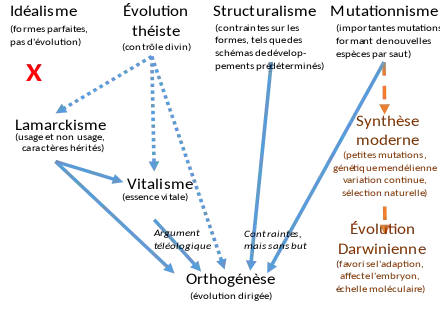
Si des explications variées de l'évolution, telles que le vitalisme, le catastrophisme et le structuralisme ont été proposées au cours du XIXe siècle, quatre alternatives importantes à la sélection naturelle étaient proposées vers 1900. L'évolution théistique prônait l'idée que Dieu pilotait directement l'évolution ; le néo-lamarckisme exposait que l'évolution était dirigée par la transmission des caractères acquis durant la vie de l'individu ; l'orthogenèse postulait que les organismes étaient affectés par des forces internes ou des lois qui dirigeaient l'évolution dans un sens particulier ; le mutationnisme affirmait que l'évolution était largement due à des mutations importantes, créant de nouvelles espèces ou formes en une seule génération.
L'évolution théistique a disparu de la littérature scientifique vers la fin du XIXe siècle quand l'appel direct à des causes surnaturelles a perdu toute crédibilité scientifique. Les autres alternatives ont compté d'importants soutiens pendant une partie du XXe siècle ; la biologie ne les a abandonnées qu'avec le développement de la génétique moderne, qui les a rendues indéfendables, et quand la génétique des populations et la théorie synthétique de l'évolution ont démontré la puissance explicative de la sélection naturelle.
Contexte
Le principe d'une modification graduelle des formes de vie sur un très grand nombre de générations (transformisme) est largement accepté dans les milieux scientifiques quelques années après la publication de L'Origine des espèces. En revanche, le fait d'accepter la sélection naturelle en tant que moteur de ce changement est beaucoup moins immédiat[1]. Six objections sont émises, au XIXe siècle, face à la théorie de Darwin[2] :
- Les séquences de fossiles sont encore fragmentaires, laissant de nombreux chaînons manquants et suggérant des sauts évolutifs[2] ;
- Le physicien Lord Kelvin calcule, en 1862, que la Terre devrait s'être complètement refroidie en 100 millions d'années ou moins à partir de sa formation, ne laissant pas assez de temps pour l'évolution — La découverte de la radioactivité résout ce problème un peu plus tard[3] - [2] — ;
- Il est avancé que de nombreuses structures sont non-fonctionnelles, et ne peuvent avoir évolué par sélection naturelle[2] ;
- Certaines structures semblent avoir évolué selon des schémas récurrents, comme les yeux des mammifères et des calmars, créatures non-apparentées[2] ;
- La capacité créative de la sélection naturelle est mise en doute, tandis que les variations entre individus sont considérées comme globalement non significatives[2] ;
- L'ingénieur Fleeming Jenkin remarque avec raison, en 1868, que l'hérédité par mélange défendue par Darwin s'oppose à l'action de la sélection naturelle. L'adoption de la génétique mendélienne au début du XXe siècle répond à ce problème, avec les notions de gènes dominants et récessifs[2] - [4] - [5].
Darwin, aussi bien que son défenseur Thomas Henry Huxley, surnommé à l'époque « le bulldog de Darwin », admettaient d'ailleurs, eux aussi, que la sélection naturelle n'était peut-être pas une solution complète ; Darwin était prêt à accepter une certaine proportion de lamarckisme, tandis qu'Huxley ne voyait pas d'inconvénient aux idées saltationnistes et orthogénétiques[6].
Au tournant du xxe siècle, la critique contre la sélection naturelle est telle qu'en 1903 le botaniste allemand Eberhard Dennert (de) écrit : « Nous observons le darwinisme sur son lit de mort » et, en 1907, Vernon Myman Lyman Kellogg, entomologiste de l'Université Stanford, défenseur de la sélection naturelle, affirmait que « le constat honnête est que la sélection darwinienne, du point de vue de sa capacité supposée à être un mécanisme explicatif indépendant et suffisant de la descendance, est aujourd'hui sérieusement discréditée dans le monde de la biologie[7] ». Il ajoute néanmoins qu'il existe des difficultés empêchant l'acceptation généralisée de l'une ou l'autre des théories alternatives, les mutations importantes étant trop rares, tandis que les mécanismes sous-jacents du lamarckisme et de l'orthogenèse manquent de preuves expérimentales[8]. Ernst Mayr écrit qu'une revue des publications et ouvrages de références de l'époque montre que, même en 1930, le point de vue faisant de la sélection naturelle le principal facteur dans l'évolution est minoritaire, seuls quelques généticiens des populations étant strictement sélectionnistes[9].
Recherche d'alternatives
La recherche de mécanismes alternatifs à la sélection naturelle est motivée par plusieurs facteurs, certains antérieurs à la parution de l'ouvrage de Darwin. La sélection naturelle, où la compétition et la mort jouent un rôle déterminant, semble à certains naturalistes immorale, elle laisse en outre peu de place pour la téléologie ou l'idée de progrès dans le développement de la vie[10] - [11]. Certains scientifiques et philosophes, comme George Jackson Mivart et Charles Lyell, qui en viennent à accepter l'évolution mais n'aiment pas le concept de sélection naturelle, soulèvent des objections religieuses[12]. D'autres, dont Herbert Spencer, le botaniste George Henslow (fils de John Stevens Henslow, botaniste et mentor de Darwin) et Samuel Butler, considèrent l'évolution comme un processus de progrès que la seule sélection naturelle ne peut expliquer. D'autres encore, comme les paléontologues américains Edward Drinker Cope et Alpheus Hyatt, ont un point de vie idéaliste et pensent que la nature, y compris le développement de la vie, suit des schémas ordonnés que la sélection naturelle ne peut expliquer[4].
Par ailleurs, un nouveau groupe de biologistes émerge à la fin du XIXe siècle, incarné par les généticiens Hugo de Vries et Thomas Hunt Morgan, lesquels veulent faire de la biologie une science expérimentale, où les résultats en laboratoire sont vitaux. Ils accordent donc peu de confiance aux travaux de naturalistes comme Darwin ou Wallace, fondés sur des observations sur le terrain de la variabilité, de l'adaptation et de la biogéographie, éléments qu'ils perçoivent comme trop anecdotiques. À la place, ils privilégient des sujets comme la physiologie et la génétique, domaines explorables par la méthode expérimentale, ce qui n'est guère le cas de la sélection naturelle et du niveau d'adaptation des organismes à leur environnement[13].
Théories anti-darwiniennes durant cette période
Évolution théiste

La science britannique se développe au début du XIXe siècle sur la base d'une théologie naturelle pour laquelle l'adaptation des espèces à leur environnement est une preuve de leur création délibérée par une volonté divine. Les concepts philosophiques empruntés à l'idéalisme allemand inspirent l'idée d'un plan ordonné présidant à la création, que Richard Owen réconcilie avec la théologie naturelle grâce au fait que l'homologie semble montrer une conception intelligente. De façon similaire, Louis Agassiz voit dans la théorie de la récapitulation le symbole d'une séquence créative dans laquelle l'humanité est l'objectif d'un plan divin. En 1844, l'ouvrage anonyme Vestiges of the Natural History of Creation, présente le concept d'Agassiz comme un évolutionnisme théistique. Robert Chambers, qui le publie anonymement, propose une loi de développement progressif, décidée par Dieu, faisant du transformisme une extension de la théorie de la récapitulation. Ce travail popularise l'idée mais est vigoureusement condamné par la communauté scientifique. Agassiz reste farouchement opposé à l'évolution et, après son départ pour les États-Unis en 1846, l'influence de son argument en faveur d'un développement ordonné s'accroit[14]. En 1858, Owen propose prudemment que ce développement pourrait être l'expression d'une loi créative continue, mais il se distancie des transformistes. Deux ans après sa critique du livre de Darwin, On the Origin of Species, Owen attaque Darwin tout en soutenant ouvertement l'évolution[15], marquant sa préférence pour un modèle de transformation obéissant à des lois. Cet argument téléologique idéaliste est repris par d'autres naturalistes, dont George Jackson Mivar et le duc d'Argyll, qui rejettent la sélection naturelle au profil d'un développement prédéterminé[16].
Parmi les soutiens de Darwin, beaucoup acceptent l'évolution, avec le postulat qu'elle peut être réconciliée avec la notion de dessein divin. Asa Gray, en particulier, tient la sélection naturelle pour le mécanisme principal de l'évolution et tente de le réconcilier avec la théologie naturelle. Il entrevoit dans la sélection naturelle une solution au problème du mal : la souffrance serait un mal nécessaire pour produire un plus grand bien, qui serait l'adaptation. Il concède que cette vision des choses a ses difficultés, et suggère que Dieu peut influencer les variations par lesquelles la sélection naturelle agit pour guider l'évolution[17]. Pour Darwin et Huxley, de telles influences surnaturelles se situent au-delà du domaine de l'investigation scientifique, et George Frederick Wright (en), un prêtre ordonné qui travailla avec Grey sur le développement de l'évolution théiste, insiste sur la nécessité de chercher des causes secondaires ou connues plutôt que d'invoquer des explications supranaturelles : « si nous cessons d'observer cette règle, c'est la fin de toute science et de toute science valable[18] ».
Une version sécularisée de cette pensée naturaliste est accueillie par une génération plus jeune de scientifiques, lesquels cherchent à étudier les causes naturelles du changement et rejettent l'évolution théiste en science. En 1872, le darwinisme dans le sens général (la factualité de l'évolution) est accepté comme point de départ. Vers 1890 seuls quelques hommes plus âgés s'attachent encore à l'idée de dessein intelligent, qui disparait du débat scientifique avant 1900. La perception des implications de la sélection naturelle reste problématique, et ceux recherchant des buts ou des finalités dans l'évolution se tournent vers le lamarckisme ou l'orthogenèse comme explications naturelles[19].
Néo-lamarckisme

Jean-Baptiste de Lamarck avait, à l'origine, proposé une théorie transformiste qui se basait largement sur une transition progressive vers une complexité accrue. Lamarck pensait aussi, comme beaucoup à son époque, que les caractères acquis au cours de la vie d'un organisme pouvaient être transmis à la génération suivante, et voyait là un mécanisme d'évolution secondaire qui produisait à une adaptation à l'environnement. Typiquement, de tels caractères concernaient les changements causés par l'usage ou l'absence d'usage d'un organe. Ce mécanisme fut, bien plus tard, connu sous le nom de lamarckisme[20]. Alors qu'Alfred Russel Wallace rejetait complètement cette idée au profit de la sélection naturelle, Charles Darwin lui-même a toujours inclus dans sa théorie ce qu'il appelait les « effets produits par la sélection naturelle sur l'accroissement de l'usage et du non-usage des parties[21] » dans On the Origin of Species, donnant pour exemple des oiseaux de grande taille se nourrissant au sol, comme l'autruche, pour laquelle l'exercice semblait avoir fortifié les jambes et atrophié les ailes, au point de rendre le vol impossible[22].
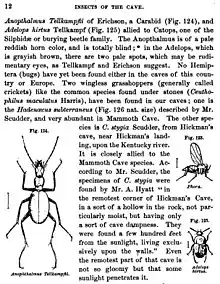
À la fin du XIXe siècle le terme de néo-lamarckisme commence à être utilisé pour désigner la position des naturalistes qui voient l'héritage de caractères acquis comme le principal mécanisme de l'évolution. Les défenseurs de cette posture sont, entre autres, l'auteur britannique et critique du darwinisme Samuel Butler, le biologiste allemand Ernst Haeckel défenseur de Darwin, les paléontologues américains Edward Drinker Cope et Alpheus Hyatt et leur compatriote, l'entomologiste Alpheus Packard. Ils voient dans le lamarckisme une idée philosophiquement supérieure à la conception darwiniste de la sélection naturelle appliquée à des variations aléatoires. Butler et Cope croient tous deux que ce changement progressif permet aux organismes de contrôler pour ainsi dire leur propre évolution, car les spécimens ayant adopté de nouveaux comportements peuvent changer le modèle d'utilisation de leurs organes, et ainsi lancer le processus évolutif. De plus, Cope et Haeckel pensent que l'évolution est un phénomène progressif. L'idée d'une progression linéaire est un élément important de la théorie de la récapitulation de Haeckel, selon laquelle le développement d'un embryon répète les grandes étapes de l'histoire évolutive de l'espèce à laquelle il appartient. Cope et Hyatt recherchent, et pensent avoir trouvé, des modèles de progressions linéaires dans les séquences fossiles[23] - [24]. Packard soutient que la perte de la vision chez les insectes troglodytes aveugles qu'il a étudié peut s'expliquer au mieux par un processus lamarckien d'atrophie par mésusage des organes, combiné avec l'hérédité des caractères acquis[25]. Packard a par ailleurs consacré un livre à Lamarck et à ses écrits[23] - [26].
Beaucoup de défenseurs américains du néo-lamarckisme sont fortement influencés par Louis Agassiz, certains comme Hyatt et Packard ayant été ses étudiants. Agassiz défend une vision idéaliste de la nature, en rapport avec la théologie naturelle qui insiste sur l'ordre et la régularité. Agassiz n'acceptera jamais l'évolution, ses disciples l'acceptent cependant, mais ils poursuivent sa recherche de schémas ordonnés dans la nature, qui leur semblent cohérents avec un ordre divin, et préfèrent les mécanismes néo-lamarckiens et orthogénétiques qui semblent plus à même de produire de tels schémas[23] - [26].
En Grande-Bretagne, le botaniste George Henslow, fils du mentor de Darwin, John Stevens Henslow, est un défenseur important du néo-lamarckisme. Il étudie comment le stress environnemental affecte le développement des plantes, et affirme que les variations ainsi provoquées peuvent en large part expliquer l'évolution. L'historien des sciences Peter Bowler écrit que, comme la plupart des Lamarckiens du XIXe siècle, Henslow ne semble pas comprendre la nécessité de démontrer que ces variations induites par certaines conditions environnementales seraient transmises à la descendance après la disparition de ces conditions, il se contente de supposer qu'il en serait ainsi[27].
Polarisation du débat : le plasma germinatif de Weismann

Les critiques du néo-lamarckisme soulèvent que personne n'a jamais produit de preuve solide de la transmission des caractères acquis. Les travaux expérimentaux du biologiste allemand August Weismann conduisent à la théorie de l'hérédité par le plasma germinatif. Cela l'amène à déclarer que l'héritage de caractères acquis est impossible, la barrière de Weismann empêchant tout changement acquis dans les cellules somatiques après la naissance d'être transmis à la prochaine génération. La conséquence est une polarisation du débat entre darwiniens et néo-lamarckistes, chacun étant obligé de choisir d'accepter ou non les idées de Weismann (considéré comme néo-darwinien ou ultra-darwinien[28]) et, par conséquent, d'admettre, ou non, la sélection naturelle comme seul moteur de l'évolution[29]. Malgré les critiques de Weismann, le néo-lamarckisme reste l'alternative à la sélection naturelle la plus défendue à la fin du XIXe siècle, et certains naturalistes s'y tiennent bien après le début du XXe siècle[24] - [30].
Effet Baldwin
Du fait du débat sur la viabilité du néo-lamarckisme, en 1896, James Mark Baldwin, Henry Fairfield Osborn et Lloyd Morgan proposent, indépendamment, un mécanisme où l'apprentissage de comportements peut causer l'évolution de nouveaux instincts ou de nouveaux traits physiques par le biais de la sélection naturelle, sans nécessiter l'héritage de caractères acquis. Leur hypothèse est la suivante : si un individu d'une espèce tire bénéfice de l'apprentissage d'un comportement nouveau, la capacité à apprendre ce comportement peut être favorisée par la sélection naturelle. Cet effet Baldwin reste un sujet de recherche et de débat dans la communauté scientifique jusqu'à nos jours[31].
Orthogenèse
L'orthogenèse est la théorie voulant que la vie aurait une tendance innée au changement, de façon linéaire, dans une direction particulière. Le terme a été popularisé par Theodor Gustav Heinrich Eimer, zoologiste allemand. Il étudie la coloration des papillons et pense avoir découvert des caractéristiques n'apportant aucun avantage en termes d'adaptation, et ne pouvant donc s'expliquer par la sélection naturelle. Eimer adhère également à l'hérédité larmarckienne des caractères acquis, mais il pense que des lois de développement interne déterminent quelles caractéristiques seraient acquises et guideraient ainsi l'évolution à long terme dans certaines directions[32].
L'orthogenèse remportant une adhésion importante au XIXe siècle, on trouve parmi ses défenseurs le biologiste russe Lev Berg et le paléontologue américain Henry Fairfield Osborn[33]. L'orthogenèse a en particulier ses défenseurs parmi certains paléontologues, qui voient dans la chronologie des fossiles des traces de changement graduel et directionnel. Ceux qui acceptent cette idée, cependant, n'adhérent pas nécessairement à un mécanisme explicatif téléologique. Ils pensent que les tendances orthogénétiques n'ont pas de valeur adaptative ; ils perçoivent, au contraire, dans certains cas, ces tendances comme préjudiciables à l'organisme étudié, comme dans les cas des bois surdimensionnés du Megaloceros giganteus qui auraient, selon eux, causé l'extinction de l'espèce[32].
Le soutien à l'orthogenèse commence à décliner avec la synthèse moderne des années 1940, où il apparaît que cette théorie ne peut rendre compte des séquences de fossiles, pour lesquelles l'analyse statistique montre un foisonnement complexe de formes apparentées. Quelques biologistes restent cependant attachés à l'idée jusqu'aux années 1950, insistant sur la différence entre microévolution et macroévolution[4] - [5].
Mutationnisme
%252C_by_Th%C3%A9r%C3%A8se_Schwartze_(1851-1918).jpg.webp)
Le mutationnisme est l'idée selon laquelle de nouvelles formes ou espèces apparaissent d'un coup, sous l'effet de mutations importantes. Il est vu comme une variante beaucoup plus rapide du concept darwinien d'un changement graduel par de petites variations aléatoires validées par la sélection naturelle. Cette idée a des défenseurs parmi les premiers généticiens comme Hugo de Vries qui, avec Carl Correns, contribue à la redécouverte des lois mendéliennes de la génétique en 1900, William Bateson, zoologiste britannique devenu généticien et, au début de sa carrière, Thomas Hunt Morgan[34] - [35].
La théorie de l'évolution par mutation, dans sa forme de 1901, défend l'idée que les espèces subissent des périodes de mutations rapides, peut-être du fait d'un stress environnemental, qui peuvent produire des mutations multiples et, dans certains cas, des espèces complètement nouvelles, en une seule génération. La théorie est d'abord proposée par le botaniste hollandais Hugo de Vries. De Vries cherche des preuves de mutations suffisamment étendues pour produire une nouvelle espèce en une seule génération et pense les avoir trouvées avec son travail de mise en culture de l'onagre du genre Oenothera, travail commencé en 1886. Les plantes avec lesquelles De Vries travaille semblent produire constamment de nouvelles variétés, avec des modifications frappantes de formes et de couleurs, dont certaines semblent être des espèces nouvelles parce que les plants de la nouvelle génération ne peuvent être croisés qu'entre eux, et non avec leurs parents. De Vries concède un rôle à la sélection naturelle pour déterminer quelles nouvelles espèces vont survivre, mais certains généticiens influencés par son travail, dont Morgan, estiment que la sélection naturelle n'est plus nécessaire du tout. Les idées de De Vries sont influentes au cours des deux premières décennies du XXe siècle, certains biologistes estimant que le mutationnisme pouvait expliquer l'apparition soudaine de nouvelles formes parmi les fossiles ; la recherche sur Oenothera se répand dans le monde entier. Cependant, les critiques, y compris de nombreux naturalistes de terrain, se demandent pourquoi aucun autre organisme ne semble présenter des mutations comparables[36].
Faiblesses des théories concurrentes
Morgan, un défenseur de la théorie des mutations de De Vries, espère trouver des preuves pour cette théorie en travaillant sur la mouche Drosophila melanogaster dans son laboratoire en 1907. Cependant, c'est un chercheur de ce même laboratoire, Hermann Joseph Muller, qui détermine en 1918 que les nouvelles variétés que De Vries avait obtenu avec Oenothera (herbe aux ânes) sont la manifestation d'hybridations polyploïdes et non le fait de mutations rapides[37] - [38]. Même s'ils doutent de l'importance de la sélection naturelle, les travaux de généticiens comme Morgan, Bateson, De Vries et d'autres apportent la preuve, entre 1900 et 1915, de la validité de la génétique mendélienne et de la théorie chromosomique de Sutton et Boveri, et valident ainsi la critique du néo-lamarckisme formulée par August Weismann en écartant l'hypothèse de l'hérédité des caractères acquis. Les travaux de l'équipe de Morgan sur les drosophiles discréditent aussi le concept d'orthogenèse en montrant le caractère aléatoire des mutations[39].
Fin de l'éclipse
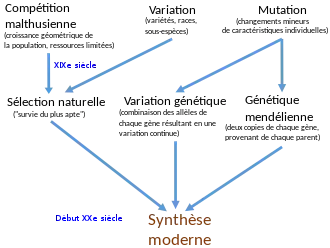
Entre 1916 et 1932, la génétique des populations doit une bonne part de son développement aux travaux de Ronald Aylmer Fisher, John Burdon Sanderson Haldane et Sewall Wright. Dans leurs travaux, ils admettent que la grande majorité des mutations produisent des effets minimes permettant d'accroître la variabilité génétique de la population, plutôt que de créer soudainement de nouvelles espèces comme le supposaient les mutationnistes. Ils commencent à produire des modèles mathématiques de la génétique d'une population prenant en compte la sélection naturelle, telle que conçue par Darwin, comme moteur de l'évolution[40]. Ernst Mayr signale que, vers 1930, la plupart des ouvrages de référence développaient encore ces mécanismes non darwiniens[9].
Les développements en génétique persuadent les naturalistes de terrain, tels Bernhard Rensch et Ernst Mayr, d'abandonner les idées néo-lamarckiennes sur l'évolution au début des années 1930[41]. À la fin de cette décennie, Ernst Mayr et Theodosius Dobjansky ont procédé à une synthèse de la génétique des populations et des connaissances de terrain sur la variabilité des populations sauvages et l'importance des populations génétiquement distinctes, particulièrement les populations séparées géographiquement ; ce fut le début de la théorie synthétique de l'évolution[42]. En 1944, George Gaylord Simpson intègre à la synthèse le domaine de la paléontologie, en analysant statistiquement les fossiles pour montrer qu'ils étaient cohérents avec une évolution créant sans cesse de multiples branches, comme prévu par la synthèse moderne, plutôt qu'avec un schéma linéaire, directionnel, attendu par les tenants du lamarckisme et de l'orthogenèse[43]. Mayr écrit qu'à la fin de ce processus, la sélection naturelle, couplée à des mécanismes aléatoires tel que la dérive génétique, était devenue l'explication universelle du changement évolutif[9].
Historiographie
Peu avant son emploi par Julian Huxley, l'expression « éclipse du darwinisme » se trouve dans un manuscrit non publié du naturaliste David Starr Jordan, dans un sens qui implique que le darwinisme est en déclin relatif, l'intérêt des biologistes se portant ailleurs[28].
La notion d'éclipse suggère une pause dans les recherches basées sur le paradigme darwinien, impliquant donc que cette période est précédée par une période d'intense activité darwinienne parmi les biologistes. Cependant, des historiens des sciences, tel Mark Largent, affirment que, même si les biologistes de l'époque acceptent majoritairement les nombreuses preuves pour l'évolution présentées dans The Origin of Species, il reste en revanche plus de réticence pour accepter la sélection naturelle comme mécanisme explicatif. Les biologistes cherchent de préférence des explications alternatives (ou complémentaires) plus compatibles avec leur vision du monde, dont des croyances selon lesquelles l'évolution doit être dirigée et constituer une forme de progrès. De plus, l'idée d'une période d'« éclipse » est commode pour des scientifiques tels que Julian Huxley, qui veulent faire apparaitre la synthèse moderne comme une nouvelle réalisation majeure, et, pour cela, gagnent à faire apparaitre la période précédente comme sombre et confuse[28].
Largent[28] estime que le livre Evolution: The Modern Synthesis, publié en 1942 par Huxley, suggère que ce qu'on appelle la synthèse moderne a commencé après une longue période d'éclipse s'étendant jusqu'aux années 1930, pendant laquelle de longues batailles opposent les mendéliens, les néo-lamarckiens, les mutationnistes et les weismanniens, mais aussi les embryologistes expérimentaux et les récapitulationnistes haeckeliens. Le concept d'éclipse permet aussi à Huxley de prendre du recul par rapport au risque d'être associé à des idées comme le darwinisme social, l'eugénisme, l'impérialisme et le militarisme[28]. Largent fait remarquer que l'historien des sciences Michael Ruse, dans son volumineux ouvrage Monad to man[44], fait l'impasse sur la quasi-totalité des biologistes américains pro-évolution du début du XXe siècle. Largent propose, comme alternative à la notion d'éclipse, le terme d'« interphase du darwinisme », en référence à l'interphase qui est une période d'inactivité apparente dans le cycle cellulaire[28].
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « The eclipse of Darwinism » (voir la liste des auteurs).
- Quammen 2006, p. 205.
- Bowler 1983, p. 23–26.
- Bowler 1983, p. 3.
- Bowler 2003, p. 196–253.
- Larson 2004, p. 105–129.
- Bowler 1983, p. 28.
- Endersby 2007, p. 143,453.
- Larson 2004, p. 128.
- Mayr et Provine 1998, p. x.
- Bowler 2003, p. 197.
- Larson 2004, p. 119–120.
- Quammen 2006, p. 209–210.
- Endersby 2007, p. 143–147, 182.
- Bowler 1983, p. 44–49.
- Secord 2001, p. 424, 512.
- Bowler 1983, p. 46, 49–50.
- Bowler 2003, p. 203–206.
- Larson 2004, p. 110–111.
- Bowler 1983, p. 26–27, 44–45, 54–55.
- Bowler 2003, p. 86–95.
- Darwin 1921, p. 159.
- Darwin 1872, p. 108.
- Bowler 2003, p. 236–244.
- Larson 2004, p. 125–129.
- Packard et Putnam 1872.
- Quammen 2006, p. 217–219.
- Bowler 2003, p. 239–240.
- Largent 2009.
- Bowler 1983, p. 41–42.
- Bowler 2003, p. 253–255.
- Bowler 2003, p. 243, 367.
- Quammen 2006, p. 221.
- Bowler 2003, p. 249.
- Bowler 2003, p. 265–270.
- Larson 2004, p. 127–129, 157–167.
- Endersby 2007, p. 148–162.
- Endersby 2007, p. 202–205.
- Ramsey et Ramsey 2014.
- Bowler 2003, p. 269–272.
- Mayr et Provine 1998, p. xi–xii.
- Mayr et Provine 1998, p. 124–127, 296.
- Mayr et Provine 1998, p. xii–xiii.
- Bowler 2003, p. 337.
- Ruse 1996.
Annexes
Bibliographie
Par ordre chronologique.
- (en) Charles Darwin, The Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life, Londres, John Murray, , 576 p. (ISBN 1-904633-78-1, lire en ligne).
- (en) Alpheus Spring Packard et Frederic Ward Putnam, The Mammoth Cave and its Inhabitants, Salem Naturalists' Agency, (OCLC 04356215, lire en ligne), p. 12.
- Charles Darwin (trad. de l'anglais par Édmond Barbier), De l'origine des espèces au moyen de la sélection naturelle ou la lutte pour l'existence dans la nature, Alfred Costes, (lire en ligne
 [PDF])
[PDF]) - (en) Julian Huxley, Evolution : the modern synthesis, Londres, Allen and Unwin, .
- (en) Peter J. Bowler, The Eclipse of Darwinism : anti-Darwinian evolutionary theories in the decades around 1900, Johns Hopkins University Press, , 291 p. (ISBN 978-0-8018-4391-4, lire en ligne).
- (en) Michael Ruse, Monad to Man : The Concept of Progress in Evolutionary Biology, Harvard University Press, , 640 p. (ISBN 978-0-674-03248-4).
- (en) Ernst Mayr et W. B. Provine, The Evolutionary synthesis : perspectives on the unification of biology, Cambridge, Mass./London, Harvard University Press, , 487 p. (ISBN 0-674-27225-0).
- (en) James A. Secord, Victorian Sensation : The Extraordinary Publication, Reception, and Secret Authorship of Vestiges of the Natural History of Creation, University of Chicago Press, , 624 p. (ISBN 978-0-226-74410-0, lire en ligne).
- (en) Peter J. Bowler, Evolution : The History of an Idea, University of California Press, , 464 p. (ISBN 0-520-23693-9, lire en ligne).
- Cédric Grimoult, Histoire de l’histoire des sciences : Historiographie de l’évolutionnisme dans le monde francophone, Librairie Droz, , 312 p. (ISBN 978-2-600-00828-0, lire en ligne).
- (en) Edward J. Larson, Evolution : The Remarkable History of Scientific Theory, Modern Library, (ISBN 0-679-64288-9).
- (en) David Quammen, The Reluctant Mr. Darwin, Atlas Books, , 304 p. (ISBN 0-393-05981-2).
- (en) Jim Endersby, A Guinea Pig's History of Biology, Cambridge (Mass.), Harvard University Press, , 499 p. (ISBN 978-0-674-02713-8 et 0-674-02713-2, lire en ligne).
- (en) Mark A. Largent, « The So-Called Eclipse of Darwinism. Descended from Darwin: Insights into the History of Evolutionary Studies, 1900–1970 », Transactions of the American Philosophical Society, American Philosophical Society, vol. 99, no 1, (JSTOR 27757422, lire en ligne [PDF]).
- (en) Justin Ramsey et Tara S. Ramsey, « Ecological studies of polyploidy in the 100 years following its discovery », Phil. Trans. Royal Society B, vol. 5, no 369, , p. 20130352 (PMID 4071525, DOI 10.1098/rstb.2013.0352, lire en ligne).
- Thierry Hoquet, Darwin contre Darwin : Comment lire 'L'Origine des espèces', Paris, Le Seuil, , 438 p. (ISBN 978-2-02-098299-3).
- Michel Morange, Une histoire de la biologie, Points, , 448 p. (ISBN 978-2-7578-7002-0).
- Jean Gayon, Darwin et l'après-Darwin : Une histoire de l'hypothèse de sélection naturelle, Paris/impr. en Pologne, Matériologiques, , 550 p. (ISBN 978-2-37361-196-0).