Virus de la rubéole
Rubella virus
| Royaume | Riboviria |
|---|---|
| Règne | Orthornavirae |
| Embranchement | Kitrinoviricota |
| Classe | Alsuviricetes |
| Ordre | Hepelivirales |
Le Virus de la rubéole, RuV, Rubella virus, est l'agent pathogène de la maladie connue sous le nom de rubéole, et il est aussi la cause de la rubéole congénitale (lorsque l'infection se produit durant les premières semaines de la grossesse). L’Homme est le seul réservoir connu de ce virus.
Le virus de la rubéole est la seule espèce du genre Rubivirus, et de la famille des Matonaviridae.
L'ARN du génome situé à l'intérieur de la capside a une longueur d'environ 9 757 nucléotides et code deux protéines non structurelles ainsi que trois protéines de structure[4]. La protéine de capside et les deux protéines d'enveloppe glycosylées, E1 et E2, constituent les trois protéines de structure.
Les bases moléculaires du mécanisme de la rubéole congénitale ne sont pas encore complètement établies, mais les études in vitro sur des lignées cellulaires ont montré que le virus de la rubéole provoque l’apoptose de certains types de cellules. Il existe des preuves d'un mécanisme P53 dépendant[5].
Taxonomie et phylogénie
Jusqu'en 2018, le genre Rubivirus était rangé dans la famille Togaviridae (Togaviridés), mais il en a depuis été sorti pour constituer le seul genre de la famille Matonaviridae (Matonaviridés). Cette nouvelle famille porte le nom de George de Maton, qui en 1814 distingua pour la première fois la rubéole de la rougeole et de la scarlatine[6]. Ce changement a été fait par le Comité international de taxonomie des virus (ICTV), l'organe directeur central de la classification virale. En raison de son génome à ARN et de son ARN polymérase ARN-dépendante, la famille Matonaviridae appartient, comme Togaviridae, à l'embranchement des Negarnaviricota.
Plusieurs raisons expliquent la séparation de Matonaviridae (Rubivirus) et Togaviridae (Alphavirus)[6] :
- morphologie : alors que les virions des alphavirus sont sphériques et contiennent une nucléocapside icosaédrique, ceux du rubivirus sont pléomorphes et n'en contiennent pas ;
- phylogénie : l'analyse phylogénétique de l'ARN polymérase ARN-dépendante des alphavirus, du virus de la rubéole et d'autres virus à ARN monocaténaire à polarité positive montre que les deux genres initialement rassemblés au sein des Togaviridés ne sont pas monophylétiques. En particulier, le virus de la rubéole est plus proche des membres des familles Benyviridae, Hepeviridae (en) et Alphatetraviridae (en), ainsi que de plusieurs virus non classés, que des membres du genre Alphavirus ;
- transmission : les alphavirus sont généralement transmis par des arthropodes, alors que le rubivirus est un virus respiratoire, transmis par les humains.
Deux virus apparentés à celui de la rubéole (RuV), qui seront sans doute rangés dans la même famille, ont été décrits en 2020[7]. Le virus Ruhugu (RuhV), le plus proche parent de RuV, a été trouvé en Ouganda chez des chauves-souris apparemment saines, des phyllorhines laineuses (en) (Hipposideros cyclops). Le virus Rustrela (RusV), proche mais extérieur au clade regroupant RuV et RuhV, a été trouvé dans un zoo allemand chez des mammifères placentaires et marsupiaux souffrant d'une encéphalite aiguë, et aussi chez des mulots à collier (Apodemus flavicollis) sauvages dans le même zoo et à proximité. Les virus Ruhugu et Rustrela ont la même architecture génomique que le virus de la rubéole, et les séquences d'acides aminés de plusieurs protéines des trois virus sont très semblables. Il apparaît que certains membres de la famille des Matonaviridés peuvent franchir des barrières interspécifiques, ce qui rend probable l'origine zoonotique du virus de la rubéole[7].
Structure
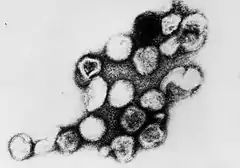
Les particules virales sphériques (virions) des Togaviridae ont un diamètre de 50 à 70 nm et sont recouvertes par une membrane lipidique (enveloppe virale), dérivée de la membrane de la cellule hôte. Il existe au premier plan des «pointes» (projections) du 6 nm composées des protéines d'enveloppe virale E1 et E2, incorporées dans la membrane[8].
L’intérieur de l'enveloppe lipidique est une capside de 40 nm de diamètre.
Réplication
Les Togavirus s’attachent à la surface des cellules via des récepteurs spécifiques et sont pris en charge par un endosome en formation. Dans les conditions de pH neutre à l'extérieur de la cellule la protéine d'enveloppe E2 recouvre la protéine E1. L’abaissement du pH au cœur de l'endosome libère la zone externe de E1 et provoque la fusion de l'enveloppe virale avec la membrane endosomale. Ensuite, la capside atteint le cytosol, se désintègre et libère le génome
Le virus (+)ssRNA, (ARN simple brin à polarité positive) agit d'abord uniquement comme une matrice pour la transcription des protéines non structurelles, qui sont synthétisées sous forme d'un grand polypeptide et sont ensuite découpées en protéines unitaires. Les séquences des protéines structurelles sont d'abord répliquées par l'ARN polymérase virale (replicase) par un fragment (-)ssRNA, un ARN complémentaire simple brin à polarité négative comme matrice et traduit séparément sous forme d’un ARNm court. Cet ARN subgénomique court est en outre enveloppé dans un virion[9].
La transcription des protéines de structure produit également un polypeptide de grande taille (110 Uma). Elle est ensuite découpée par protéolyse en E1, E2 et protéine de capside. E1 et E2 sont des protéines transmembranaires de type I qui sont transportées dans le réticulum endoplasmique avec l'aide d'une séquence signal amino-terminale. À partir du réticulum le complexe hétérodimérique E2 E1 atteint l’appareil de Golgi, où se produit le bourgeonnement de nouveaux virions (à la différence des virus alpha, où la prolifération a lieu au niveau de la membrane plasmique). Les protéines de capside, d'autre part restent dans le cytoplasme et interagissent avec l'ARN génomique, pour former la capside[10].
Protéine de capside
La protéine de capside (protéine C) a des fonctions différentes. Ses principales missions sont la formation de l'homooligomère pour former la capside, et la liaison avec l'ARN génomique. En outre, elle est responsable de l'agrégation de l'ARN dans la capside, interagit avec les protéines membranaires E1 et E2 et se lie à la protéine p32 de l'hôte humain qui a un rôle important pour la réplication du virus à l’intérieur de l'hôte[11].
Contrairement à l'alpha virus, la capside ne subit pas d’autoprotolyse, mais elle est coupée du reste du polypeptide par la peptidase. La production de la capside se produit à la surface des membranes intracellulaires en même temps que le bourgeonnement du virus[12].
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Rubella virus » (voir la liste des auteurs).
- ICTV. International Committee on Taxonomy of Viruses. Taxonomy history. Published on the Internet https://talk.ictvonline.org/., consulté le 28 février 2021
- ICTV. International Committee on Taxonomy of Viruses. Taxonomy history. Published on the Internet https://talk.ictvonline.org/., consulté le 28 février 2021
- ICTV. International Committee on Taxonomy of Viruses. Taxonomy history. Published on the Internet https://talk.ictvonline.org/., consulté le 28 février 2021
- (en) Dominguez G, Wang CY, Frey TK, « Sequence of the genome RNA of rubella virus: evidence for genetic rearrangement during togavirus evolution », Virology, vol. 177, no 1, , p. 225–38 (PMID 2353453)
- (en) Megyeri K, Berencsi K, Halazonetis TD, et al, « Involvement of a p53-dependent pathway in rubella virus-induced apoptosis », Virology, vol. 259, no 1, , p. 74–84 (PMID 10364491, DOI 10.1006/viro.1999.9757)
- Rubing Chen, Suchetana Mukhopadhyay, Andres Merits, Bethany Bolling, Farooq Nasar et al., « Create a new family Matonaviridae to include the genus Rubivirus, removed from the family Togaviridae », sur ICTV (consulté le ).
- (en) Andrew J. Bennett, Adrian C. Paskey, Arnt Ebinger, Florian Pfaff, Grit Priemer et al., « Relatives of rubella virus in diverse mammals », Nature, (DOI /10.1038/s41586-020-2812-9).
- (en) Bardeletti G, Kessler N, Aymard-Henry M, « Morphology, biochemical analysis and neuraminidase activity of rubella virus », Arch. Virol., vol. 49, nos 2-3, , p. 175–86 (PMID 1212096)
- « Togaviridae- Classification and Taxonomy »
- (en) Garbutt M, Law LM, Chan H, Hobman TC, « Role of rubella virus glycoprotein domains in assembly of virus-like particles », J. Virol., vol. 73, no 5, , p. 3524–33 (PMID 10196241, PMCID 104124, lire en ligne)
- (en) Beatch MD, Everitt JC, Law LJ, Hobman TC, « Interactions between rubella virus capsid and host protein p32 are important for virus replication », J. Virol., vol. 79, no 16, , p. 10807–20 (PMID 16051872, PMCID 1182682, DOI 10.1128/JVI.79.16.10807-10820.2005)
- (en) Beatch MD, Hobman TC, « Rubella virus capsid associates with host cell protein p32 and localizes to mitochondria », J. Virol., vol. 74, no 12, , p. 5569–76 (PMID 10823864, PMCID 112044, lire en ligne)
Références biologiques
- (en) Référence ICTV : Matonaviridae (consulté le )
- (en) Référence ICTV : Rubivirus (consulté le )
- (en) Référence ICTV : Rubella virus (consulté le )
Voir aussi
Bibliographie
- David M. Knipe, Peter M. Howley et al. (eds.): Fields´ Virology 4. Auflage, Philadelphia 2001
- C.M. Fauquet, M.A. Mayo et al.: Eighth Report of the International Committee on Taxonomy of Viruses, London San Diego 2005
Liens externes
- (en) Viralzone: Rubivirus