Viroporine
Une viroporine est une petite protéine virale multifonctionnelle, généralement hydrophobe, qui modifie les membranes plasmiques et facilite de ce fait la libération des virions par les cellules infectées[2] - [3]. Les viroporines sont capables de s'assembler en canaux ioniques ou pores oligomériques dans la membrane plasmique de la cellule hôte, ce qui rend cette membrane plus perméable. De nombreuses viroporines présentent par ailleurs d'autres effets sur le métabolisme cellulaire et l'homéostasie faisant intervenir des interactions protéine-protéine avec des protéines de la cellule hôte[3]. Elles ne sont pas nécessairement indispensables à la réplication virale mais la favorisent sensiblement. On les trouve dans une variété de génomes mais sont particulièrement présentes dans les virus à ARN. De nombreux virus à l'origine de maladies humaines expriment des virporines, comme le virus de l'hépatite C , le virus de l'immunodéficience humaine (VIH-1), le virus de la grippe A, le poliovirus, le virus respiratoire syncytial et le coronavirus du syndrome respiratoire aigu sévère (SARS-CoV)[3] - [4] - [5].
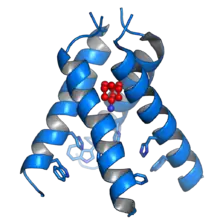
Structure
Les viroporines sont généralement des protéines de 100 à 120 résidus d'acides aminés et contiennent au moins un domaine capable de se replier en hélice transmembranaire amphiphile. Certaines contiennent des acides aminés à chaîne latérale basique ou des segments à résidus aromatiques dont on pense qu'ils sont localisés à l'interface avec la membrane[3]. Des oligomères de ces protéines, le plus souvent des tétramères[6] , forment des canaux ioniques ou des pores à sélectivité ionique généralement faible qui permet la diffusion des ions à travers la membrane plasmique. L'architecture moléculaire du pore, sa sélectivité ionique, son niveau d'intégration dans la membrane cellulaire et son extension éventuelle de part et d'autre de cette membrane sont des paramètres très variables d'une viroporine à l'autre, ce qui indique que ces protéines ont des rôles fonctionnels diversifiés[4] - [5].
Classification
Il est possible de classer les viroporines en quatre classes en fonction de leur topologie et de leur orientation dans la membrane. Les viroporines de classe I on une seule hélice transmembranaire tandis que celles de classe II présentent un motif hélice-coude-hélice dans lequel les deux hélices traversent la membrane. La sous-classe IA correspond aux viroporines à une seule hélice transmembranaire dont l'extrémité C-terminale est située côté cytosol tandis que la sous-classe IB correspond aux viroporines orientées dans l'autre sens, avec l'extrémité N-terminale côté cytosol. De même, la sous-classe IIA correspond aux viroporines dont le coude est orienté vers le cytosol et les deux extrémités peptidiques sont orientés hors de celui-ci — vers le milieu extracellulaire dans le cas d'une membrane plasmique ou vers le lumen du réticulum endoplasmique ou d'une vésicule dans le cas du système endomembranaire — tandis que la sous-classe IIB correspond aux viroporines orientées en sens inverse[5]. Il existe cependant des exceptions à ce schéma, comme la protéine non structurelle 4 (NSP4 (en)) des rotavirus[7] - [8].
Notes et références
- (en) Amanda L. Stouffer, Rudresh Acharya, David Salom, Anna S. Levine, Luigi Di Costanzo, Cinque S. Soto, Valentina Tereshko, Vikas Nanda, Steven Stayrook et William F. DeGrado, « Structural basis for the function and inhibition of an influenza virus proton channel », Nature, vol. 451, no 7178, , p. 596-599 (PMID 18235504, DOI 10.1038/nature06528, Bibcode 2008Natur.451..596S, lire en ligne)
- (en) Luis Carrasco, « Modification of Membrane Permeability by Animal Viruses », Advances in Virus Research, vol. 45, , p. 61-112 (PMID 7793329, DOI 10.1016/S0065-3527(08)60058-5, lire en ligne)
- (en) Maria Eugenia Gonzalez et Luis Carrasco, « Viroporins », FEBS Letters, vol. 552, no 1, , p. 28-34 (PMID 12972148, DOI 10.1016/S0014-5793(03)00780-4, lire en ligne)
- (en) Jose L. Nieto-Torres, Carmina Verdiá-Báguena, Carlos Castaño-Rodriguez, Vicente M. Aguilella et Luis Enjuanes, « Relevance of Viroporin Ion Channel Activity on Viral Replication and Pathogenesis », Viruses, vol. 7, no 7, , p. 3552-3573 (PMID 26151305, PMCID 4517115, DOI 10.3390/v7072786, lire en ligne)
- (en) José Luis Nieva, Vanesa Madan et Luis Carrasco, « Viroporins: structure and biological functions », Nature Reviews Microbiology, vol. 10, no 8, , p. 563-574 (PMID 22751485, DOI 10.1038/nrmicro2820, lire en ligne)
- (en) Kai Wang, Shiqi Xie et Bing Sun, « Viral proteins function as ion channels », Biochimica et Biophysica Acta (BBA) - Biomembranes, vol. 1808, no 2, , p. 510-515 (PMID 20478263, DOI 10.1016/j.bbamem.2010.05.006, lire en ligne)
- (en) José Luis Nieva et Luis Carrasco, « Viroporins: Structures and functions beyond cell membrane permeabilization », Viruses, vol. 7, no 10, , p. 5169-5171 (PMID 26702461, PMCID 4632374, DOI 10.3390/v7102866, lire en ligne)
- (en) Liya Hu, Sue E. Crawford, Joseph M. Hyser, Mary K. Estes et B. V. Venkataram Prasad, « Rotavirus non-structural proteins: structure and function », Current Opinion in Virology, vol. 2, no 4, , p. 380-388 (PMID 22789743, PMCID 3422752, DOI 10.1016/j.coviro.2012.06.003, lire en ligne)