Haplogroupe I-M253
L'haplogroupe I-M253, aussi connu comme I1, est un haplogroupe du chromosome Y. Les marqueurs génétiques identifiants confirmés de I-M253 sont les SNP's M253,M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186, et L187. C'est une branche primaire de l'Haplogroupe I-M170 (I*).

| Date d'origine | 3170–5070 av. J.-C. (précédemment 11000 av. J.-C. à 33000 av. J.-C.) |
|---|---|
| Place d'origine | Europe du Nord |
| Ancêtre | I* (M170) |
| Descendants |
I1a (DF29/S438); I1b (S249/Z131); I1c (Y18119/Z17925) |
| Mutations définies | M253, M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186, L187 |
L'haplogroupe atteint ses fréquences de pointe en Suède (52 % des hommes dans le comté de Västra Götaland) et à l'ouest de la Finlande (plus de 50 % dans la province de Satakunta)[1] En termes de moyennes nationales, I-M253 se situe entre 35 et 38 % des hommes suédois[2], 32,8 % des hommes danois, environ 31,5 % des mâles norvégiens[3] et environ 28 % des hommes finlandais.
L'haplogroupe I-M253 est une branche primaire de l'haplogroupe I* (I-M170), qui a été présente en Europe depuis l'antiquité. L'autre branche primaire de I* est-I-M438, également connue comme I2.
Avant un reclassement en 2008[4], le groupe était connu comme I1a, un nom qui a depuis été réaffecté à une branche primaire, l'haplogroupe I-DF29. Les autres principales branches de I1 (M253) sont I1b (S249/Z131) et I1c (Y18119/Z17925).
Origines
Selon une étude publiée en 2010, I-M253 est apparu entre 3 170 et 5 000 ans, au Chalcolithique en Europe[5]. Une nouvelle étude en 2015 estime son origine entre 3 470 et 5 070 ans ou entre 3 180 et 3 760, à l'aide de deux techniques différentes[6]. Il est suggéré qu'il a initialement essaimé à partir de la région qui est maintenant le Danemark[7].
Une étude de 2014 en Hongrie révéla les restes de neuf personnes de la culture rubanée, dont un portait le SNP M253 qui définit l'haplogroupe I1. Cette culture perdure entre 5 500 à 4 700 ans av. J.-C.[8].
Structure
I-M253 (M253, M307.2/P203.2, M450/S109, P30, P40, L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157.1, L186, et L187) ou I1 [9];
- I-DF29 (DF29/S438); I1a
- I-CTS6364 (CTS6364/Z2336); I1a1
- I-M227; I1a1a
- I-L22 (L22/S142); I1a1b
- I-P109; I1a1b1
- I-L205 (L205.1/L939.1/S239.1); I1a1b2
- I-Z74; I1a1b3
- I-L300 (L300/S241); I1a1b4
- I-L287
- I-L258 (L258/S335)
- I-L813
- I-L287
- I-Z58 (S244/Z58); I1a2
- I-Z59 (S246/Z59); I1a2a
- I-Z60 (S337/Z60, S439/Z61, Z62); I1a2a1
- I-Z140 (Z140, Z141)
- I-L338
- I-F2642 (F2642)
- I-Z73
- I-L1302
- I-L573
- I-L803
- I-Z140 (Z140, Z141)
- I-Z382; I1a2a2
- I-Z60 (S337/Z60, S439/Z61, Z62); I1a2a1
- I-Z138 (S296/Z138, Z139); I1a2b
- I-Z2541
- I-Z59 (S246/Z59); I1a2a
- I-Z63 (S243/Z63); I1a3
- I-BY151; I1a3a
- I-L849.2; I1a3a1
- I-BY351; I1a3a2
- I-CTS10345
- I-Y10994
- I-Y7075
- I-CTS10345
- I-S2078
- I-S2077
- I-Y2245 (Y2245/PR683)
- I-L1237
- I-FGC9550
- I-S10360
- I-S15301
- I-Y7234
- I-L1237
- I-Y2245 (Y2245/PR683)
- I-S2077
- I-BY62 (BY62); I1a3a3
- I-BY151; I1a3a
- I-CTS6364 (CTS6364/Z2336); I1a1
- I-Z131 (Z131/S249); I1b
- I-CTS6397; I1b1
- I-Z17943 (Y18119/Z17925, S2304/Z17937); I1c
Répartition géographique
I-M253 est trouvé à densité élevée dans le Nord de l'Europe et d'autres pays qui ont subi d'importantes migrations de l'Europe du Nord, soit dans la Période de Migration, la période Viking de ou de l'époque moderne. On le trouve dans tous les endroits envahis par les anciens Germains et les Vikings.
Au cours de l'ère moderne, d'importants populations I-M253 ont d'ailleurs pris racine dans des pays d'immigration et anciennes colonies européennes telles que les États-Unis, l'Australie et le Canada.
| Population | Taille de l'échantillon | I (total) | I1 (I-M253) | I1a1a (I-M227) | Source |
|---|---|---|---|---|---|
| Autriche | 43 | 9.3 | 2.3 | 0.0 | Underhill et al. 2007 |
| Biélorussie: Vitebsk | 100 | 15 | 1.0 | 0.0 | Underhill et al. 2007 |
| Biélorussie: Brest | 97 | 20.6 | 1.0 | 0.0 | Underhill et al. 2007 |
| Bosnie | 100 | 42 | 2.0 | 0.0 | Rootsi et al. 2004 |
| Bulgarie | 808 | 26.6 | 4.3 | 0.0 | Karachanak et al. 2013 |
| République Tchèque | 47 | 31.9 | 8.5 | 0.0 | Underhill et al. 2007 |
| République Tchèque | 53 | 17.0 | 1.9 | 0.0 | Rootsi et al. 2004 |
| Danemark | 122 | 39.3 | 32.8 | 0.0 | Underhill et al. 2007 |
| Angleterre | 104 | 19.2 | 15.4 | 0.0 | Underhill et al. 2007 |
| Estonie | 210 | 18.6 | 14.8 | 0.5 | Rootsi et al. 2004 |
| Estonie | 118 | 11.9 | Lappalainen et al. 2008 | ||
| Finlande (national) | 28.0 | Lappalainen et al. 2006 | |||
| Finlande: ouest | 230 | 40 | Lappalainen et al. 2008 | ||
| Finlande: est | 306 | 19 | Lappalainen et al. 2008 | ||
| Finlande: région de Satakunta | 50+ | Lappalainen et al. 20089 | |||
| France | 58 | 17.2 | 8.6 | 1.7 | Underhill et al. 2007 |
| France | 12 | 16.7 | 16.7 | 0.0 | Cann et al. 2002 |
| France (Basse-Normandie) | 42 | 21.4 | 11.9 | 0.0 | Rootsi et al. 2004 |
| Allemagne | 125 | 24 | 15.2 | 0.0 | Underhill et al. 2007 |
| Grèce | 171 | 15.8 | 2.3 | 0.0 | Underhill et al. 2007 |
| Hongrie | 113 | 25.7 | 13.3 | 0.0 | Rootsi et al. 2004 |
| Irlande | 100 | 11 | 6.0 | 0.0 | Underhill et al. 2007 |
| Tatars de Kazan | 53 | 13.2 | 11.3 | 0.0 | Trofimova 2015 |
| Lettonie | 113 | 3.5 | Lappalainen et al. 2008 | ||
| Lituanie | 164 | 4.9 | Lappalainen et al. 2008 | ||
| Pays-Bas | 93 | 20.4 | 14 | 0.0 | Underhill et al. 2007 |
| Norvège | 2826 | 31.5 | Eupedia 2017 | ||
| Russie (national) | 16 | 25 | 12.5 | 0.0 | Cann et al. 2002 |
| Russie: Pskov | 130 | 16.9 | 5.4 | 0.0 | Underhill et al. 2007 |
| Russie: Kostroma | 53 | 26.4 | 11.3 | 0.0 | Underhill et al. 2007 |
| Russie: Smolensk | 103 | 12.6 | 1.9 | 0.0 | Underhill et al. 2007 |
| Russie: Voronez | 96 | 19.8 | 3.1 | 0.0 | Underhill et al. 2007 |
| Russie: Arkhangelsk | 145 | 15.8 | 7.6 | 0.0 | Underhill et al. 2007 |
| Russie: Cossaques | 89 | 24.7 | 4.5 | 0.0 | Underhill et al. 2007 |
| Russie: Caréliens | 140 | 10 | 8.6 | 0.0 | Underhill et al. 2007 |
| Russie: Caréliens | 132 | 15.2 | Lappalainen et al. 2008 | ||
| Russie: Vepsa | 39 | 5.1 | 2.6 | 0.0 | Underhill et al. 2007 |
| Slovaquie | 70 | 14.3 | 4.3 | 0.0 | Rootsi et al. 2004 |
| Slovénie | 95 | 26.3 | 7.4 | 0.0 | Underhill et al. 2007 |
| Suède (national) | 160 | 35.6 | Lappalainen et al. 2008 | ||
| Suède (national) | 38.0 | Lappalainen et al. 2009 | |||
| Suède: Västra Götaland | 52 | Lappalainen et al. 2009 | |||
| Suisse | 144 | 7.6 | 5.6 | 0.0 | Rootsi et al. 2004 |
| Turquie | 523 | 5.4 | 1.1 | 0.0 | Underhill et al. 2007 |
| Ukraine: Lvov | 101 | 23.8 | 4.9 | 0.0 | Underhill et al. 2007 |
| Ukraine: Ivanovo-Frankov | 56 | 21.4 | 1.8 | 0.0 | Underhill et al. 2007 |
| Ukraine: Hmelnitz | 176 | 26.2 | 6.1 | 0.0 | Underhill et al. 2007 |
| Ukraine: Cherkassy | 114 | 28.1 | 4.3 | 0.0 | Underhill et al. 2007 |
| Ukraine: Belgorod | 56 | 26.8 | 5.3 | 0.0 | Underhill et al. 2007 |
Grande-Bretagne
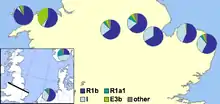
En 2002, un document a été publié par Michael E. Weale et collègues en montrant la preuve génétique des différences entre la population anglaise et galloise, y compris un niveau nettement plus élevé de l'ADN-Y de l'haplogroupe I en Angleterre que dans le pays de Galles. Ils ont vu cela comme une preuve convaincante de l'invasion massive anglo-saxonne de l'est de la Grande-Bretagne à partir de l'Allemagne du nord et du Danemark au cours de la Période de Migration[10]. Les auteurs ont supposé que les populations avec des grandes proportions d'haplogroupe I furent originaires de l'Allemagne du nord ou du sud de la Scandinavie, en particulier du Danemark, et que leurs ancêtres avaient migré à travers la Mer du Nord avec les migrations des Anglo-Saxons et les Vikings danois. La principale revendication faite par les rechercheurs était:
qu'un événement d'immigration anglo-saxonne affectant de 50 à 100 % du fonds génétique des hommes de l'Angleterre centrale serait nécessaire à l'époque. Nous observons, toutefois, que nos données ne nous permettent pas de distinguer un événement simplement ajouté au fonds génétique masculin indigène de l'Angleterre centrale, d'un autre où les populations de mâles autochtones étaient déplacés ailleurs, ou bien où le nombre d'hommes autochtones a été réduit ... Cette étude montre que la frontière galloise était plus une barrière génétique aux flux de gènes du chromosome Y anglo-saxon que la Mer du Nord. Ces résultats indiquent qu'une frontière politique peut être plus important qu'une géophysique dans la structuration génétique des populations.
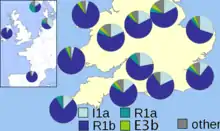
En 2003, un document publié par Christian Capelli et les collègues prit en charge, mais modifié, les conclusions de Weale et collègues[11]. Ce document, qui a échantillonné sur une grille la Grande-Bretagne et l'Irlande , a trouvé une petite différence entre les échantillons gallois et anglais, avec une diminution progressive de fréquence de l'haplogroupe I en se déplaçant vers l'ouest dans le sud de la Grande-Bretagne. Les résultats suggérèrent aux auteurs que les envahisseurs vikings norvégiens avaient fortement influencé le secteur nord des Îles Britanniques, mais que les deux échantillons anglais et le écossais (de l’île principale) ont tous les deux de l'influence allemande/danoise .
Membres éminents de l'I-M253
Alexander Hamilton, par la généalogie et les tests de ses descendants (en supposant être la paternité réelle correspondante de sa généalogie), a été placé dans l'haplogroupe ADN-Y I-M253[12].
Birger Jarl, "Duc de Suède" de la Maison des Goths de Bjalbo, fondateur de Stockholm, dont les vestiges enfouis dans une église avaient été faits tester en 2002 et confirmés aussi I-M253
Les Passagers du Mayflower William Brewster, Edward Winslow et George Soule grâce à des tests ADN
Marqueurs
Voici les spécifications techniques pour les mutations SNP et STR de I'haplogroupe I-M253.
Nom: M253[13]
- Type: SNP
- Source: M (Peter Underhill de « l'Université de Stanford »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ))
- Position: « ChrY:13532101..13532101 (+ verticale) »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le )
- Position (paires de bases): 283
- Taille totale (paires de bases): 400
- Longueur: 1
- ISOGG HG: I1
- Amorce F (sense 5'→ 3'): GCAACAATGAGGGTTTTTTTG
- Amorce R (anti-sense 5'→ 3'): CAGCTCCACCTCTATGCAGTTT
- YCC HG: I1
- Changement de nucléotides allèles (mutation): C à T
Nom: M307[14]
- Type: SNP
- Source: M (Peter Underhill)
- Position: « ChrY:21160339..21160339 (+ verticale) »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le )
- Longueur: 1
- ISOGG HG: I1
- Amorce F: TTATTGGCATTTCAGGAAGTG
- Amorce R: GGGTGAGGCAGGAAAATAGC
- YCC HG: I1
- Changement de nucléotides allèles (mutation): G pour A
Nom: P30[15]
- Type: SNP
- Source: PS (« Michael Hammer »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ) de l' Université de l'Arizona et James F. Wilson, de l'Université d'Édimbourg)
- Position: « ChrY:13006761..13006761 (+ verticale) »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le )
- Longueur: 1
- ISOGG HG: I1
- Amorce F: GGTGGGCTGTTTGAAAAAGA
- Amorce R: AGCCAAATACCAGTCGTCAC
- YCC HG: I1
- Changement de nucléotides allèles (mutation): G pour A
- Région: ARSDP
Nom: P40[16]
- Type: SNP
- Source: PS (Michael Hammer et James F. Wilson)
- Position: « ChrY:12994402..12994402 (+ verticale) »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le )
- Longueur: 1
- ISOGG HG: I1
- Amorce F: GGAGAAAAGGTGAGAAACC
- Amorce R: GGACAAGGGGCAGATT
- YCC HG: I1
- De nucléotides allèles changement (mutation): C à T
- Région: ARSDP
Références
- T. Lappalainen, V. Laitinen, E. Salmela et P. Andersen, « Migration Waves to the Baltic Sea Region », Annals of Human Genetics, vol. 72, no 3, , p. 337–348 (PMID 18294359, DOI 10.1111/j.1469-1809.2007.00429.x).
- T. Lappalainen, U. Hannelius, E. Salmela et U. von Döbeln, « Population Structure in Contemporary Sweden: A Y-Chromosomal and Mitochondrial DNA Analysis », Annals of Human Genetics, vol. 73, no 1, , p. 61–73 (PMID 19040656, DOI 10.1111/j.1469-1809.2008.00487.x).
- (en) Eupedia, Distribution of European Y-chromosome DNA (Y-DNA) haplogroups by country in percentage, .
- Tatiana M. Karafet, F. L. Mendez, M. B. Meilerman et P. A. Underhill, « New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree », Genome Research, vol. 18, no 5, , p. 830–8 (PMID 18385274, PMCID 2336805, DOI 10.1101/gr.7172008).
- (en) Pedro Soares, Alessandro Achilli, Ornella Semino, William Davies, Vincent Macaulay, Hans-Jürgen Bandelt, Antonio Torroni, and Martin B. Richards, The Archaeogenetics of Europe, Current Biology, vol. 20 (February 23, 2010), R174–R183. yDNA Haplogroup I: Subclade I1, Family Tree DNA.
- (en) « TMRCAs of major haplogroups in Europe estimated using two methods. : Large-scale recent expansion of European patrilineages shown by population resequencing : Nature Communications : Nature Publishing Group », sur www.nature.com (consulté le ).
- (en) Peter A. Underhill et al., New Phylogenetic Relationships for Y-chromosome Haplogroup I: Reappraising its Phylogeography and Prehistory, in Rethinking the Human Revolution (2007), p. 33-42.
- (en) Anna Szécsényi-Nagy et al., « Tracing the genetic origin of Europe's first farmers reveals insights into their social organization », 2015 apr 22.
- ISOGG, Y-DNA Haplogroup I and its Subclades - 2017 (31 January 2017).
- Michael E. Weale, Deborah A. Weiss, Rolf F. Jager et Neil Bradman, « Y chromosome Evidence for Anglo-Saxon Mass Migration », Molecular Biology and Evolution, vol. 19, no 7, , p. 1008–1021 (PMID 12082121, DOI 10.1093/oxfordjournals.molbev.a004160, lire en ligne).
- Cristian Capelli, Nicola Redhead, Julia K. Abernethy et Fiona Gratrix, « A Y Chromosome Census of the British Isles », Current Biology, vol. 13, no 11, , p. 979–984 (PMID 12781138, DOI 10.1016/S0960-9822(03)00373-7, lire en ligne [PDF]).
- « Founding Father DNA », isogg.org, sur isogg.org.
- snpdev, « Reference SNP (refSNP) Cluster Report: rs9341296 », nih.gov, sur nih.gov.
- snpdev, « Reference SNP (refSNP) Cluster Report: rs13447354 », nih.gov, sur nih.gov.
- « P30 »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ).
- « P40 »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) (consulté le ).
Projets
- Haplogroupe I bases de données
- Haplogroup I1 Project at FTDNA
- Danish Demes Regional DNA Project at FTDNA
- Haplogroup I-P109 Project
- British Isles DNA Project
- Bases de données générales ADN-Y
Il existe plusieurs bases de données publiques mettant en vedette I-M253, y compris: