Ceratonykus
Ceratonykus oculatus

Ceratonykus (signifiant « griffe cornue ») est un genre éteint de dinosaures Alvarezsauridae de Mongolie qui a vécu pendant le Crétacé supérieur (stade Campanien tardif, 72,1 Ma) dans ce qui est maintenant la formation Barun Goyot. Le type et la seule espèce, Ceratonykus oculatus, est connu à partir d'un squelette fragmentaire, y compris un crâne incomplet, d'un individu adulte. Elle a été nommée et décrite en 2009 par Vladimir Alifanov et Rinchen Barsbold. Ses descripteurs ont remis en question le classement traditionnel des Alvarezsauridae dans les Theropoda, suggérant plutôt qu'ils étaient des Ornithischia, mais cela n'a pas été accepté depuis. Ceratonykus a une longueur estimée à 75 centimètres et un poids de 760 grammes. Il a été considéré comme un synonyme junior possible de Parvicursor.
Découverte et dénomination
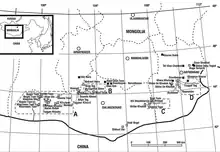
En 2003, un squelette partiel d'un Alvarezsauridae a été découvert dans la formation Barun Goyot de la localité de Khermiin Tsav, en Mongolie, par le préparateur Otkhoon Zhargal du Centre paléontologique de l'Académie des sciences de Mongolie (MPC). Le squelette était enfermé dans deux morceaux de roche au moment de la découverte. Le spécimen a ensuite été nommé et décrit en 2009 par Vladimir Alifanov et Rinchen Barsbold. Le spécimen holotype, MPC no. 100/124, consiste en un crâne et une mâchoire inférieure incomplets, des vertèbres cervicales, des vertèbres caudales, un sternum, des coracoïdes gauche et droite, un fragment d'humérus droit, des éléments distaux de membres antérieurs, un ilium fragmentaire et des membres postérieurs. Le spécimen est celui d'un individu adulte d'après la fusion des astragales et des tibias, avec des sutures partiellement oblitérées[1].
Le nom générique, Ceratonykus, est dérivé des mots grecs « ceratinus » (cornu) et « onyx » (griffe). Le nom spécifique, oculatus, est dérivé du mot latin « oculatus » (myope, gros yeux)[1].
Description
Taille et traits distinctifs
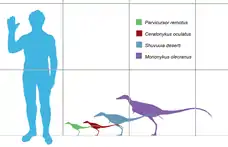
En 2016, Gregory S. Paul a estimé la longueur de Ceratonykus à 0,6 mètre et son poids à 1 kilogramme[2]. Cependant, Rubén Molina-Pérez et Asier Larramendi (2016) ont estimé la longueur de Ceratonykus à 75 centimètres et son poids à 760 grammes[3].
Alifanov & Barsbold (2009) ont initialement décrit Ceratonykus en se basant sur la longue région du crâne pré-orbitaire ; une fenestration temporale supérieure ovale qui était 0,4 aussi long que les frontaux ; la longueur du frontal étant presque quatre fois plus grande que sa largeur ; les frontaux se rétrécissant vers le rostre en un coin étroit ; les préfrontaux se rejoignant vers le milieu ; les processus basiptérygoïdes (protubérance de la base de l’os sphénoïde) étant deux tiers aussi hauts que les carrés ; les dentaires forment une profonde dépression qui s'amincit vers le rostre ; des fenêtres mandibulaires étendues ; des centrums étroits des vertèbres cervicales et caudales antérieures ; la crête deltopectorale étant séparée de la tête humérale par une encoche ; la phalange basale du doigt majeur de la main est allongée, avec ses côtés modérément larges, et un condyle distal étroit et symétrique ; la plaque postacétabulaire de l'iliaque possédant une crête craniomédiale longitudinale relativement petite ; un fémur fortement incurvé qui est presque deux fois moins long que le tibiotarse ; un quatrième trochanter distinct ; une crête craniomédiale des tibias non développée ; un processus ascendant de l'astragale qui est haut et large ; les tarso-métatarses étant 1,33 fois plus longs que les fémurs ; les deuxième et quatrième métatarses sont étroitement contigus sur toute leur étendue ; une profonde encoche est formée proximo-dorsalement entre les métatarses ; les deuxièmes métatarses sont plus courts que les quatrièmes distalement ; les tarso-métatarses sont 3,5 fois plus longs que les troisièmes métatarses ; et la phalange basale du quatrième doigt des pattes postérieures n'est que légèrement plus courte que la phalange basale du deuxième doigt[1].
Crâne
La partie du crâne la plus proche du rostre est étroite et étendue, avec une région occipitale profonde et des orbites larges. Les fenêtres anté-orbitales sont au moins deux fois moins grandes que les orbites et les barres temporales sont relativement petites, tandis que les fenêtres temporales inférieures sont étroites et légèrement plus bas en hauteur que le diamètre des orbites. Les deux frontaux sont appariés, cependant, le frontal gauche est dépourvu de la marge postéro-médiale tandis que le frontal droit n'est connu que par son extrémité rostrale. Les deux frontaux sont quatre fois plus longs que larges, comme chez Shuvuuia, et sont en forme de coin étroit en vue de face. Le squamosal jouxte le post-orbitaire et le processus paroccipital, tout en étant relativement courts et en forme de T. Les os postorbitaires ont un grand processus médian et sont reliés aux os pariétaux et frontaux par un processus différent. Le processus ventral des os lacrymaux est étroit, le processus dorsal étant très long et en forme de tige. Les jugaux et les quadratojugaux sont circulaires en section transversale et sont fusionnés mais ne forment pas de suture. Les quadratojugaux sont hauts, le condyle distal étant présent sur la crête occipitale et la crête latérale étant reliée à la face supérieure de la zone de contact des os squamosal et postorbital. Comme chez Shuvuuia, une encoche est présente entre les condyles latéraux et occipitaux. Les processus paroccipitaux de la boîte crânienne sont comprimés vers la face supérieure et la face inférieure. L'intérieur du prootique (os situé entre la base du crâne et l'oreille) présente une structure à cellules grossières, qui n'est visible que sur le côté droit du crâne. Le basioccipital forme un condyle occipital sur un cou long et large, tandis que les exoccipitaux ne font pas partie du condyle. Contrairement à Shuvuuia, les processus basiptérygoïdes divergent à presque 60° et sont presque deux tiers aussi longs que les os carrés[1].
Les dentaires sont longs, l'extrémité rostrale s'élargissant légèrement verticalement et s'incurvant vers le bas. Les fenêtres mandibulaires ont une forme ovale large et un bord antérieur qui est en ligne avec la frontière entre le premier et le deuxième tiers des orbites. Les branches de la mâchoire inférieure forment les processus para-articulaires, qui sont en ligne avec l'articulation carré-mandibulaire[1].
La base du processus occipital droit contenait une cavité. Une structure à cellules grossières est visible sur le côté droit du crâne. Le corps du basioccipital forme un condyle occipital, tandis que les exoccipitaux ne font pas partie du condyle. Les tubercules sphénoccipitaux sont séparés par un large sillon et une dépression est présente dans la zone de contact du basioccipital et du basisphénoïde. Le basisphénoïde coïncide avec les processus basiptérygoïdes, qui se compriment légèrement vers les côtés et le milieu. Les hémisphères cérébraux constituent la majorité du cerveau. Ces hémisphères cérébraux sont séparés du mésencéphale par la présence d'un sulcus (sillon) et conservent des traces de la membrane cérébrale. Un sulcus inter-hémisphérique est présent au début des voies olfactives. Les lobes optiques sont déplacés vers l'avant, ce qui est une caractéristique des oiseaux existants. Les globes oculaires auraient été positionnés vers le cerveau et ont un diamètre d'un quart de la longueur de la tête, une caractéristique qui suggère que la vue des Alvarezsauridae était le système afférent clé. Les tubercules acoustiques sont inhabituellement larges et sont positionnés entre le bord occipital des hémisphères cérébraux et le cervelet[1].
Post-crânien
L'atlas est probablement représenté par l'intercentrum, qui est bas et allongé, la région postérieure étant brisée. Un os placé verticalement est présent sur le fragment et représente probablement l'arc neural de l'atlas. La série de trois vertèbres cervicales est dépourvue de pleurocéle et possède des condyles très petits et hémisphériques. Ces condyles sont également petits dans les autres vertèbres. Les centres vertébraux s'étendent un peu plus large que ceux de Mononykus et sont un peu plus étroits que ceux de Shuvuuia. Les diapophyses des deux premières vertèbres de la série sont relativement petites mais sont plus larges à la base et s'étendent vers les côtés et la face inférieure. Les parapophyses sont séparées les unes des autres par un sillon ventral et sont reliées aux diapophyses par une crête étroite sur la vertèbre médiane. L'arc neural de cette vertèbre est large et bas, l'épine neurale étant totalement absente. La première vertèbre caudale libre a un centrum étroit et moyennement haut, avec une petite quille ventrale présente près du condyle. Tout comme la première vertèbre caudale libre, les trois vertèbres caudales antérieures articulées ont un centrum étroit mais sont hautes sur toute leur étendue. Contrairement à la première vertèbre caudale libre et aux trois vertèbres caudales antérieures articulées, deux vertèbres isolées de la région caudale moyenne ont un centre bas et une crête à peine visible à la place de l'épine neurale. Cependant, toutes les vertèbres caudales sont soudées[1].
Le sternum de la ceinture pectorale est un os non apparié qui ne présente pas de quille médiane prononcée. La surface latérale des coracoïdes est convexe, le bord postéro-dorsal étant épaissi et une courte crête se formant près du bord postéro-supérieur. La base de la tête humérale était inclinée caudalement. La crête deltopectorale était séparée de la tête humérale par une encoche, la région supérieure ressemblant à un processus élevé d'après le contour arrondi de la base et le foramen au centre. Ceratonykus a une phalange basale du doigt majeur qui est similaire à celle de Mononykus, mais diffère par la partie distale plus étroite. Le deuxième carpométacarpe possède deux os en forme de pointe de taille variable qui pourraient être des phalanges terminales de doigts plutôt. La plaque postérieure de l'ilium gauche est large, le bord latéral étant arqué et la partie craniomédiale ayant une crête longitudinale[1].
Les fémurs sont dépourvus d'épiphyses et sont convexes dans leur partie proximale, tandis que la surface médiale est droite. Les tibias manquent également d'épiphyses et les tibiotarses sont légèrement courbés vers les côtés dans la moitié supérieure. Les tibias ont une section transversale subcirculaire et sont dépourvus d'une partie saillante de la crête cnémiale externe, l'astragale étant complètement soudé aux os. Les métatarses sont une fois et demie plus longs que les fémurs, le tarsométatarse étant trois fois et demie plus long que le troisième métatarse. Des rainures dorsales et palmaires sont présentes dans la zone de connexion des métatarsiens et du tarsométatarsien, mais disparaissent dans le tiers palmaire inférieur du tarsométatarsien. Comme chez Parvicursor, les deuxièmes métatarses étaient plus courts que les quatrièmes métatarses, les métatarses étant élargis dans leur partie médiane en vue latérale. En raison de la mauvaise conservation du spécimen holotype, la forme des facettes articulaires des métatarses et des doigts n'est pas connue. On a trouvé en articulation naturelle les phalanges basales des deuxième et troisième doigts du pied droit. Ceratonykus présente un rapport de longueurs des phalanges basales similaire à celui de Mononykus : la plus grande phalange basale du quatrième doigt est légèrement plus courte que la phalange basale du deuxième doigt, et la phalange basale du troisième doigt est légèrement plus longue que la phalange basale du deuxième doigt[1].
Classification
Alifanov & Barsbold (2009) ont initialement placé Ceratonykus au sein des Alvarezsauridae, dans le clade Parvicursoridae, comme taxon frère de Mononykus. Les auteurs ont remis en question le placement traditionnel des Alvarezsauridae au sein de Theropoda, suggérant plutôt qu'ils étaient des Ornithischia, mais depuis lors, cela n'a pas été accepté[1]. Qin et al. (2019) ont retrouvé Ceratonykus au sein d'une polytomie avec Linhenykus, Shuvuuia, Mononykus et Parvicursor, une position également retrouvée par Xu et al. (2018)[4] - [5], Lee et al. (2019) l'ont retrouvé dans une position similaire, mais avec l'inclusion de Nemegtonykus et un clade contenant Kol, Xixianykus et Albinykus[6], De plus, il a également été retrouvé au sein d'une polytomie par Fowler et al. (2020), avec l'inclusion de Trierarchuncus, et Averianov & Lopatin (2021), avec l'inclusion de Khulsanurus, dans un arbre de consensus strict utilisant des caractères non pondérés[7] - [8]. Cependant, un arbre de consensus strict avec pondération implicite par Averianov & Lopatin (2021) le place comme taxon frère de Parvicursor[8]. Agnolin et al. (2012) ont positionné Ceratonykus au sein de la tribu des Ceratonykini (retenu par PBDB), comme taxon frère de Xixianykus et Albinykus[9]. Averianov & Lopatin (2022b) ont considéré Ceratonykus comme un synonyme junior de Parvicursor car les différences entre les deux taxons étaient minimes et concernaient des caractères probablement ontogénétiques et peut-être aussi des variations individuelles[10].
Une analyse phylogénétique réalisée par Averianov & Lopatin (2022b) est reproduite ci-dessous[10] :
Les résultats d'une analyse antérieure d'Agnolin et al. (2012) sont reproduits ci-dessous[9] :
| Alvarezsauridae |
| |||||||||||||||||||||||||||||||||
Paléobiologie
En 2011, Vladimir Alifanov et Sergey Savelyev ont décrit le moulage naturel du cerveau de Ceratonykus. Les auteurs ont noté que l'anatomie du cerveau est une structure linéaire, sans coudes dans le plan parasagittal aux limites entre le mésencéphale et le myélencéphale, une caractéristique qui n'est pas observée chez les crocodiles, les oiseaux et les autres dinosaures. Les analyseurs visuel et auditif sont représentés par des lobes séparés dans le mésencéphale, les lobes optiques étant déplacés ventrolatéralement comme chez les oiseaux. Alifanov et Savelyev ont testé l'hypothèse d'ancêtres grimpeurs des Alvarezsauria en examinant le cervelet de Ceratonykus, qui possède au moins un sulcus transversal comme chez les crocodiles mais pas chez les lézards. Les crocodiles ayant une meilleure coordination des mouvements que les lézards, puisqu'ils doivent s'orienter dans un espace tridimensionnel, les auteurs ont proposé que l'ancêtre des Alvarezsauridae était probablement arboricole. Les auteurs ont également suggéré que le cervelet complexe aurait pu être utile pour une régulation efficace de la vitesse et de nombreux mouvements tels que le saut chez les Alvarezsauridae terrestres. En outre, ils ont considéré que la spécialisation de la vue, de l'ouïe et de l'odorat aurait pu rendre les Alvarezsauridae efficaces pour la recherche de nourriture et la chasse[11]. Agnolin et al. (2021) ont également noté que l'élargissement de la moelle épinière du cou et l'expansion du cervelet de Ceratonykus suggèrent une amélioration locomotrice chez les Alvarezsauridae[12].
Paléoenvironnement

Ceratonykus est connu de la formation Barun Goyot, qui a été datée du stade Campanien du Crétacé supérieur. La formation de Barun Goyot a une épaisseur de 110 mètres et recouvre la formation de Djadokhta. Les sédiments de la formation ont été déposés dans des conditions diverses, la partie inférieure étant constituée d'une alternance de dépôts dunaires et de lacs qui existaient dans les zones interdunaires, tandis que la partie supérieure était constituée de sédiments déposés sur une zone similaire aux takyrs qui était inondée à intervalles irréguliers. Dans l'ensemble, la formation avait un climat semi-aride avec des précipitations importantes[13].
La faune de la formation de Barun Goyot comprend les Alvarezsauridae Khulsanurus[8], Ondogurvel[10] et Parvicursor[14], les Ankylosauridae Saichania[15], Tarchia[15], Zaraapelta[16] et un Ankylosauridae indéterminé[17] ; les Ceratopsia Bagaceratops[18] - [19] - [20], Breviceratops[19] - [21] et peut-être Udanoceratops[22] ; les Dromaeosauridae Velociraptorinae Kuru[23] et Shri[24] ; l'Unenlagiidae Halszkaraptorinae Hulsanpes[25] ; le Pachycephalosauridae Tylocephale[26] - [27] ; les Oviraptoridae Conchoraptor[28], Heyuannia[28] et Nemegtomaia[29], le Sauropoda Titanosauria Quaesitosaurus[30] ; les Enantiornithinae Gobipteryx[31] et Gobipipus[32] ; et l'oiseau Ornithuromorpha Hollanda[33].
Notes et références
Notes
Références
- V. R. Alifanov et R. Barsbold, « Ceratonykus oculatus gen. et sp. nov., a new dinosaur (?Theropoda, Alvarezsauria) from the Late Cretaceous of Mongolia », Paleontological Journal, vol. 43, no 1, , p. 94–106 (ISSN 0031-0301 et 1555-6174, DOI 10.1134/s0031030109010109, lire en ligne, consulté le )
- (en) The Princeton Field Guide to Dinosaurs, (ISBN 978-0-691-16766-4, lire en ligne)
- Rubén Molina-Pérez et Asier Larramendi, Récords y curiosidades de los dinosaurios: terópodos y otros dinosauromorfos, Larousse, (ISBN 978-84-16641-15-4, lire en ligne)
- Zichuan Qin, James Clark, Jonah Choiniere et Xing Xu, « A new alvarezsaurian theropod from the Upper Jurassic Shishugou Formation of western China », Scientific Reports, vol. 9, no 1, (ISSN 2045-2322, DOI 10.1038/s41598-019-48148-7, lire en ligne, consulté le )
- Xing Xu, Jonah Choiniere, Qingwei Tan et Roger B.J. Benson, « Two Early Cretaceous Fossils Document Transitional Stages in Alvarezsaurian Dinosaur Evolution », Current Biology, vol. 28, no 17, , p. 2853–2860.e3 (ISSN 0960-9822, DOI 10.1016/j.cub.2018.07.057, lire en ligne, consulté le )
- Sungjin Lee, Jin-Young Park, Yuong-Nam Lee et Su-Hwan Kim, « A new alvarezsaurid dinosaur from the Nemegt Formation of Mongolia », Scientific Reports, vol. 9, no 1, (ISSN 2045-2322, DOI 10.1038/s41598-019-52021-y, lire en ligne, consulté le )
- Denver W. Fowler, John P. Wilson, Elizabeth A. Freedman Fowler et Christopher R. Noto, « Trierarchuncus prairiensis gen. et sp. nov., the last alvarezsaurid: Hell Creek Formation (uppermost Maastrichtian), Montana », Cretaceous Research, vol. 116, , p. 104560 (ISSN 0195-6671, DOI 10.1016/j.cretres.2020.104560, lire en ligne, consulté le )
- Alexander O. Averianov et Alexey V. Lopatin, « The second taxon of alvarezsaurid theropod dinosaurs from the Late Cretaceous Khulsan locality in Gobi Desert, Mongolia », Historical Biology, , p. 1–12 (ISSN 0891-2963 et 1029-2381, DOI 10.1080/08912963.2021.2000976, lire en ligne, consulté le )
- Federico L. Agnolin, Jaime E. Powell, Fernando E. Novas et Martin Kundrát, « New alvarezsaurid (Dinosauria, Theropoda) from uppermost Cretaceous of north-western Patagonia with associated eggs », Cretaceous Research, vol. 35, , p. 33–56 (ISSN 0195-6671, DOI 10.1016/j.cretres.2011.11.014, lire en ligne, consulté le )
- Alexander O. Averianov et Alexey V. Lopatin, « A re-appraisal of Parvicursor remotus from the Late Cretaceous of Mongolia: implications for the phylogeny and taxonomy of alvarezsaurid theropod dinosaurs », Journal of Systematic Palaeontology, vol. 19, no 16, , p. 1097–1128 (ISSN 1477-2019 et 1478-0941, DOI 10.1080/14772019.2021.2013965, lire en ligne, consulté le )
- V. R. Alifanov et S. V. Saveliev, « Brain structure and neurobiology of Alvarezsaurians (Dinosauria), exemplified by Ceratonykus oculatus (Parvicursoridae) from the Late Cretaceous of Mongolia », Paleontological Journal, vol. 45, no 2, , p. 183–190 (ISSN 0031-0301 et 1555-6174, DOI 10.1134/s0031030111020031, lire en ligne, consulté le )
- Federico L. Agnolín, Jun-Chang Lu, Martin Kundrát et Li Xu, « Alvarezsaurid osteology: new data on cranial anatomy », Historical Biology, vol. 34, no 3, , p. 443–452 (ISSN 0891-2963 et 1029-2381, DOI 10.1080/08912963.2021.1929203, lire en ligne, consulté le )
- (en) Ryszard Gradzinski et Tomasz Jerzyklewicz, « Sedimentation of the Barun Goyot Formation », Palaeontologia Polonica, no 30, (lire en ligne [archive], consulté le )
- Karhu, A.A. and Rautian, A.S. (1996). "A new family of Maniraptora (Dinosauria: Saurischia) from the Late Cretaceous of Mongolia". Paleontological Journal Russian Academy of Sciences 30(5): 583-592.
- T. Maryańska, « Ankylosauridae (Dinosauria) from Mongolia », Palaeontologia Polonica, vol. 37, , p. 85–151
- Victoria M. Arbour, Philip J. Currie et Demchig Badamgarav, « The ankylosaurid dinosaurs of the Upper Cretaceous Baruungoyot and Nemegt formations of Mongolia », Zoological Journal of the Linnean Society, vol. 172, no 3, , p. 631–652 (ISSN 0024-4082, DOI 10.1111/zoj.12185, lire en ligne, consulté le )
- Jin-Young Park, Yuong-Nam Lee, Philip J. Currie et Michael J. Ryan, « A new ankylosaurid skeleton from the Upper Cretaceous Baruungoyot Formation of Mongolia: its implications for ankylosaurid postcranial evolution », Scientific Reports, vol. 11, no 1, (ISSN 2045-2322, DOI 10.1038/s41598-021-83568-4, lire en ligne, consulté le )
- (en) Teresa Maryańska et Halszka Osmólska, « Protoceratopsidae (Dinosauria) of Asia », Palaeontologia Polonica, no 33, (lire en ligne [archive], consulté le )
- Łukasz Czepiński, « Ontogeny and variation of a protoceratopsid dinosaur Bagaceratops rozhdestvenskyi from the Late Cretaceous of the Gobi Desert », Historical Biology, vol. 32, no 10, , p. 1394–1421 (ISSN 0891-2963 et 1029-2381, DOI 10.1080/08912963.2019.1593404, lire en ligne, consulté le )
- (en) Bitnara Kim, Hyesu Yun et Yuong-Nam Lee, « The postcranial skeleton of Bagaceratops (Ornithischia: Neoceratopsia) from the Baruungoyot Formation (Upper Cretaceous) in Hermiin Tsav of southwestern Gobi, Mongolia », sur www.jgsk.or.kr, Journal of the Geological Society of Korea, (consulté le )
- S. M. Kurzanov, « A new Late Cretaceous protoceratopsid genus from Mongolia », Paleontological Journal, vol. 24, , p. 85−91
- (en) V. S. Tereschenko, « Adaptive features of protoceratopoids (Ornithischia: Neoceratopsia) », Paleontological Journal, vol. 42, no 3, , p. 273–286 (ISSN 1555-6174, DOI 10.1134/S003103010803009X, lire en ligne, consulté le )
- James G. Napoli, Alexander A. Ruebenstahl, Bhart-Anjan S. Bhullar et Alan H. Turner, « A New Dromaeosaurid (Dinosauria: Coelurosauria) from Khulsan, Central Mongolia », American Museum Novitates, vol. 2021, no 3982, (ISSN 0003-0082, DOI 10.1206/3982.1, lire en ligne [archive], consulté le )
- Alan H. Turner, Shaena Montanari et Mark A. Norell, « A New Dromaeosaurid from the Late Cretaceous Khulsan Locality of Mongolia », American Museum Novitates, vol. 2020, no 3965, (ISSN 0003-0082, DOI 10.1206/3965.1, lire en ligne, consulté le )
- Andrea Cau et Daniel Madzia, « Redescription and affinities ofHulsanpes perlei(Dinosauria, Theropoda) from the Upper Cretaceous of Mongolia », PeerJ, vol. 6, , e4868 (ISSN 2167-8359, DOI 10.7717/peerj.4868, lire en ligne, consulté le )
- Susannah Maidment et Paul Barrett, « Osteological correlates for quadrupedality in ornithischian dinosaurs », Acta Palaeontologica Polonica, (ISSN 0567-7920, DOI 10.4202/app.2012.0065, lire en ligne [archive], consulté le )
- Robert M. Sullivan, « A taxonomic review of the Pachycephalosauridae (Dinosauria: Ornithischia) », New Mexico Museum of Natural History and Science Bulletin, no 35, , p. 347-365 (lire en ligne [archive], consulté le )
- G.F. Funston, S.E. Mendonca, P.J. Currie et R. Barsbold, « Oviraptorosaur anatomy, diversity and ecology in the Nemegt Basin », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 494, , p. 101–120 (ISSN 0031-0182, DOI 10.1016/j.palaeo.2017.10.023, lire en ligne, consulté le )
- Federico Fanti, Philip J. Currie et Demchig Badamgarav, « New Specimens of Nemegtomaia from the Baruungoyot and Nemegt Formations (Late Cretaceous) of Mongolia », PLoS ONE, vol. 7, no 2, , e31330 (ISSN 1932-6203, DOI 10.1371/journal.pone.0031330, lire en ligne, consulté le )
- (en) S.M. Kurzanov et A. F. Bannikov, « A new sauropod from the Upper Cretaceous of Mongolia », Paleontological Journal, (lire en ligne, consulté le )
- (en) Andrzej Elzanowski, « Skulls of Gobipteryx (Aves) from the Upper Cretaceous of Mongolia », Palaeontologia Polonica, no 37, (lire en ligne, consulté le )
- E. N. Kurochkin, S. Chatterjee et K. E. Mikhailov, « An embryonic enantiornithine bird and associated eggs from the cretaceous of Mongolia », Paleontological Journal, vol. 47, no 11, , p. 1252–1269 (ISSN 0031-0301 et 1555-6174, DOI 10.1134/s0031030113110087, lire en ligne, consulté le )
- Alyssa K. Bell, Luis M. Chiappe, Gregory M. Erickson et Shigeru Suzuki, « Description and ecologic analysis of Hollanda luceria, a Late Cretaceous bird from the Gobi Desert (Mongolia) », Cretaceous Research, vol. 31, no 1, , p. 16–26 (ISSN 0195-6671, DOI 10.1016/j.cretres.2009.09.001, lire en ligne, consulté le )
Annexes
Articles connexes
Références taxinomiques
- (en) Référence Paleobiology Database : Ceratonykus, Alifanov & Barsbold, 2009