Évolution dirigée
L'évolution dirigée (ED) est une méthode utilisée en ingénierie des protéines qui imite le processus de sélection naturelle pour "diriger" l'évolution des protéines ou des acides nucléiques vers un certain objectif défini par l'utilisateur[1]. Cette méthode consiste à soumettre un gène à des cycles itératifs de mutagénèse (ce qui crée une banque de variants), de sélection (qui permet d'exprimer des variants et d'isoler des membres possédant la fonction que l'on recherche) et d'amplification (ce qui génère un modèle pour le cycle suivant). Elle peut être mise en place in vivo (dans des cellules vivantes), ou in vitro (cellules libres en solution ou dans des microgouttes). L'évolution dirigée est utilisée à la fois en ingénierie des protéines comme alternative à la conception rationnelle des protéines modifiées, ainsi que dans les études des principes évolutifs fondamentaux en environnement contrôlé en laboratoire.
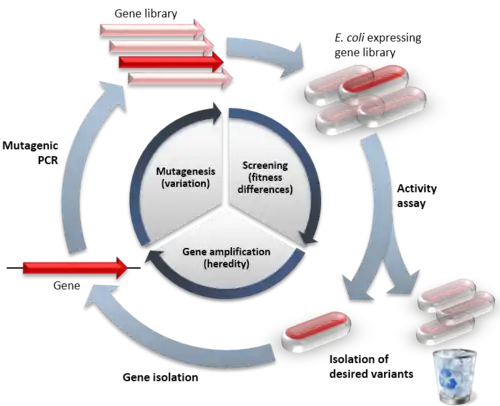
Principes
L'évolution dirigée est une imitation du cycle naturel de l'évolution dans le cadre d'un dispositif en laboratoire. L'évolution requiert trois choses pour avoir lieu : qu'il y ait de la variation (diversité génétique) entre les réplicats, que cette variation cause des différences dans la fitness sur laquelle la sélection agit, et enfin que cette variation soit héritable. En ED, un seul gène subit le processus d'évolution par des cycles itératifs de mutagénèse, de sélection ou dépistage, et d'amplification[2]. Typiquement, les cycles de ces étapes sont répétés, en utilisant le meilleur variant d'un cycle comme modèle pour le suivant afin d'améliorer le processus étape par étape.
La probabilité de succès dans une expérience d'évolution dirigée est directement liée à la taille totale de la banque, car le fait de prendre en compte plus de mutants augmente les chances d'en trouver un possédant les propriétés d'intérêt[3].
Générer de la diversité
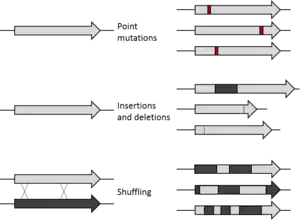
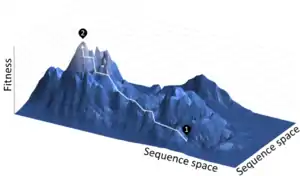
La première étape dans la réalisation d'un cycle d'ED est la création d'une banque de gènes variants. L'espace séquentiel d'une séquence aléatoire est vaste (10130 séquences possibles pour une protéine de 100 acides aminés) et extrêmement peu peuplé par des protéines fonctionnelles. Ni l'évolution expérimentale[4] ni l'évolution naturelle[5] ne peuvent approcher le fait de pouvoir échantillonner autant de séquences. Bien sûr, l'évolution naturelle échantillonne des séquences variantes proches des séquences de protéines fonctionnelles, et cela est limité en ED par la mutagénèse d'un gène déjà fonctionnel. Certains calculs suggèrent que c'est entièrement faisable dans tous les buts pratiques (fonctionnels et structurels), l'espace séquentiel protéique ayant été entièrement exploré au cours de l'évolution de la vie sur Terre[5].
Le gène de départ peut subir la mutagénèse par des mutations ponctuelles (en utilisant des mutagènes chimiques ou la PCR sujette à erreurs)[6] - [7] ainsi que par des insertions et délétions (grâce à des transposons)[8]. La recombinaison génétique peut être imitée par un remaniement de l'ADN[9] - [10] de plusieurs séquences (habituellement de plus de 70% d'homologie) afin de sauter dans des régions d'espace séquentiel entre les gènes parents remaniés. Enfin, des régions spécifiques d'un gène peuvent être systématiquement randomisées (subir une mutagénèse aléatoire)[11] pour une approche plus ciblée basée sur la connaissance de la structure et de la fonction. En fonction de la méthode utilisée, la proportion de variants fonctionnels variera d'une banque à l'autre. Même si un organisme est utilisé pour exprimer un gène d'intérêt, si seulement ce gène subit une mutagénèse, le reste du génome de l'organisme restera le même et peut être ignoré pour l'expérience (dans la mesure où on garde un environnement génétique constant).
Détecter des différences dans la fitness
La majorité des mutations sont délétères et donc les banques de mutants tendent à avoir majoritairement des variants avec une activité réduite[12]. Ainsi, une analyse à haut débit est vitale pour mesurer l'activité afin de trouver les rares variants ayant des mutations bénéfiques qui améliorent les propriétés désirées. Deux catégories principales de méthodes existent pour isoler les variants fonctionnels. Les systèmes de sélection couplent directement la fonction protéique à la survie du gène, alors que les systèmes de dépistage analysent individuellement chaque variant et permettent de fixer un seuil quantitatif pour isoler un variant ou une population de variants d'une activité désirée. À la fois la sélection et le dépistage peuvent être réalisés dans les cellules vivantes (évolution in vivo) ou directement sur la protéine ou l'ARN sans que ce soit à l'intérieur de cellules (évolution in vitro)[13] - [14].
Pendant l'évolution in vivo, chaque cellule (habituellement chez les bactéries ou les levures) est transformée avec un plasmide contenant un membre différent de la banque de variants. Dans cette méthode, seul le gène d'intérêt diffère entre les cellules, tous les autres gènes restant les mêmes. Les cellules expriment la protéine soit dans leur cytoplasme soit à leur surface, où leur fonction peut être testée. Ce format a l'avantage de sélectionner des propriétés dans un environnement cellulaire, ce qui est utile lorsque la protéine ou l'ARN évolué(e) est utilisé(e) dans des organismes vivants. Dans la méthode sans cellules, l'ED implique l'utilisation de transcription-traduction in vitro pour produire des protéines ou de l'ARN libre(s) en solution ou compartimentalisé(es) dans des microgouttes artificielles. Cette méthode a l'avantage d'être plus versatile dans les conditions de sélection (par exemple la température, le solvant), et peut exprimer des protéines qui peuvent être toxiques pour les cellules. De plus, les expériences d'évolution in vitro peuvent générer des banques beaucoup plus grandes (jusqu'à 1015) parce que la banque d'ADN n'a pas besoin d'être insérée dans les cellules (ce qui constitue souvent une étape limitante).
Sélection
Sélectionner l'activité de liaison est conceptuellement simple. La molécule cible est immobilisée sur un support solide, on fait couler une banque de protéines variantes sur ce support, celles qui se lient faiblement sont emportées et les variants liants restants sont récupérés afin d'isoler leurs gènes[15]. La liaison d'une enzyme à un inhibiteur covalent immobilisé a également été utilisée pour tenter d'isoler des catalyseurs actifs. Cependant, cette approche sélectionne seulement un turnover catalytique unique et ne constitue pas un bon modèle de liaison au substrat ou une bonne réactivité au substrat. Si une activité enzymatique peut se révéler nécessaire pour la survie de la cellule, soit par la synthèse d'un métabolite vital soit par destruction d'une toxine, alors la survie de la cellule est fonction de l'activité enzymatique[16] - [17]. De tels systèmes sont généralement limités en débit seulement par la transformation efficace des cellules. Ils sont également moins chers et exigeants en main-d'œuvre que le dépistage, néanmoins ils sont typiquement difficiles à concevoir, enclins aux artéfacts et ne donnent aucune information sur la gamme d'activités présentes dans la banque.
Dépistage
Une alternative à la sélection est le système de dépistage. Chaque gène variant est exprimé individuellement et analysé pour mesurer quantitativement l'activité (le plus souvent grâce à un indicateur coloré ou un fluorochrome). Les variants sont ensuite classés et l'expérimentateur décide quels variants utiliser comme modèles pour le cycle suivant d'évolution dirigée. Même les analyses avec le plus haut débit ont une couverture moins large que les méthodes de sélection, mais ont l'avantage de donner des informations détaillées sur chacun des variants dépistés. Ces données désagrégées peuvent aussi être utilisées afin de caractériser la distribution des activités dans les banques, ce qui n'est pas possible dans les simples systèmes de sélection. Ainsi, les systèmes de dépistage apportent des avantages certains lorsqu'il s'agit de caractériser expérimentalement l'évolution adaptative et les paysages adaptatifs.
Assurer l'hérédité

Lorsque les protéines fonctionnelles ont été isolées, il est nécessaire que leurs gènes le soient aussi ; en conséquence, un lien génotype-phénotype est requis[16]. Ce lien peut être covalent, comme dans l'affichage d'ARNm où le gène de l'ARNm est lié à la protéine par la puromycine à la fin de la traduction[4]. Autrement, la protéine et son gène peuvent être co-localisés par compartimentalisation dans les cellules vivantes[18] ou dans des gouttelettes d'émulsion[19]. Les séquences géniques isolées sont ensuite amplifiées par PCR ou par des bactéries-hôtes transformées. On peut utiliser soit la meilleure séquence, soit un groupe de séquences, en tant que modèles pour le cycle suivant de mutagénèse. Les cycles répétés de diversification-sélection-amplification génèrent des variants protéiques adaptés aux pressions de sélection appliquées.
Comparaison avec la conception rationnelle des protéines
Avantages de l'évolution dirigée
La conception rationnelle d'une protéine est basée sur une connaissance approfondie de la structure de cette protéine, ainsi que de son mécanisme catalytique[20] - [21]. Des changements spécifiques sont ensuite effectués via une mutagénèse dirigée dans le but de changer la fonction de la protéine. L'inconvénient de cela est que même quand la structure et le mécanisme d'action de la protéine sont bien connus, le changement dû à la mutation est encore difficile à prédire. Ainsi, un avantage de l'ED est qu'il n'y a pas besoin de comprendre le mécanisme de l'activité désirée ou de comment les mutations l'affecteraient[22].
Limites de l'évolution dirigée
Une restriction de l'ED est qu'une analyse à haut débit est requise afin de mesurer les effets d'un grand nombre de différentes mutations aléatoires. Cela peut demander une recherche et un développement extensifs avant qu'on puisse l'utiliser pour l'ED. De plus, de telles analyses sont souvent très spécifiques à l'observation d'une activité particulière, et ne sont donc pas transférables à de nouvelles expériences d'ED[23].
De plus, sélectionner une amélioration dans la fonction analysée génère simplement des améliorations dans cette fonction. Pour comprendre comment ces améliorations sont atteintes, les propriétés de l'enzyme qui évolue doivent être mesurées. Une amélioration de l'activité analysée peut être due à des améliorations dans l'activité catalytique de l'enzyme ou dans la concentration de cette enzyme. Cela ne confère également pas de garantie qu'une amélioration dans un substrat va améliorer l'activité dans un autre. Cela est particulièrement important lorsque l'activité désirée ne peut pas être directement dépistée ou sélectionnée, et on utilise donc un substrat "proxy". L'ED peut mener à une spécialisation évolutive du proxy sans améliorer l'activité désirée. En conséquence, choisir un dépistage ou des conditions de sélections approprié(e)s est vital pour qu'une ED réussisse.
Approches combinatoires
Les chercheurs étudient des approches combinées "semi-rationnelles" afin de répondre aux limites de la conception rationnelle et l'évolution dirigée[1] - [24]. Les mutations bénéfiques sont rares, et donc des nombres élevés de mutants aléatoires doivent subir un dépistage afin de trouver des variants améliorés. Les "banques ciblées" se concentrent sur des régions randomisées que l'on suppose plus riches en mutations bénéfiques pour l'étape de mutagénèse de l'ED. Une banque ciblée contient moins de variants qu'une banque de mutants aléatoires traditionnelle et donc n'a pas besoin d'une telle analyse à haut débit.
Le fait de créer une banque ciblée requiert une certaine connaissance des résidus à muter dans la structure ; par exemple, la connaissance du site actif d'une enzyme peut permettre seulement aux résidus connus pour interagir avec le substrat d'être randomisés[25] - [26]. Autrement, la connaissance de quelles régions protéiques sont variables dans la nature peut guider la mutagénèse seulement dans ces régions-là[27] - [28].
Utilisations
L'évolution dirigée est fréquemment utilisée en ingénierie des protéines comme une alternative à la conception rationnelle[29], mais peut aussi être utilisée pour étudier des questions fondamentales d'évolution des enzymes[30].
Ingénierie des protéines
En tant qu'outil d'ingénierie des protéines, l'ED a eu le plus de succès dans trois domaines :
- Améliorer la stabilité des protéines à utilisation biotechnologique, à des hautes températures ou dans des solvants puissants[31] - [32] ;
- Améliorer l'affinité de liaison d'anticorps thérapeutiques (maturation de l'affinité)[33] et l'activité d'enzymes conçues de novo[22] ;
- Modifier la spécificité de substrat d'enzymes existantes[34] - [35] - [36] - [37] (souvent destinées à être utilisées dans l'industrie)[29].
Études évolutives
L'étude de l'évolution naturelle est traditionnellement basée sur des organismes encore existants de nos jours, et leurs gènes. Cependant, la recherche est fondamentalement limitée par le manque de fossiles (et particulièrement le manque de séquences d'ADN fossile)[38] - [39] et une connaissance incomplète des anciennes conditions environnementales. L'évolution dirigée aborde l'évolution dans un système contrôlé de gènes codant des enzymes individuelles[40] - [41] - [24] - [37], des ribozymes[42] et des réplicateurs[43] - [44] (voir la théorie du gène égoïste de Richard Dawkins), de façon similaire à l'évolution expérimentale des eucaryotes[45] - [46], procaryotes[47] et virus[48].
L'ED permet de contrôler la pression de sélection, le taux de mutation et l'environnement (à la fois au niveau des facteurs abiotiques comme la température, et des facteurs biotiques comme les autres gènes présents dans l'organisme). De plus, il existe un dossier complet listant tous les gènes évolutifs intermédiaires. Cela permet d'avoir des mesures détaillées des processus évolutifs, par exemple l'épistasie, l'évolvabilité, les paysages adaptatifs[49] contraints dans l'adaptationnisme[50] - [51], et les réseaux neutres[52].
Voir aussi
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Directed evolution » (voir la liste des auteurs).
Références
- Stefan Lutz, « Beyond directed evolution--semi-rational protein engineering and design », Current Opinion in Biotechnology, vol. 21, no 6, , p. 734–743 (ISSN 1879-0429, PMID 20869867, PMCID PMC2982887, DOI 10.1016/j.copbio.2010.08.011, lire en ligne, consulté le )
- C. A. Voigt, S. Kauffman et Z. G. Wang, « Rational evolutionary design: the theory of in vitro protein evolution », Advances in Protein Chemistry, vol. 55, , p. 79–160 (ISSN 0065-3233, PMID 11050933, lire en ligne, consulté le )
- Paul A. Dalby, « Strategy and success for the directed evolution of enzymes », Current Opinion in Structural Biology, vol. 21, no 4, , p. 473–480 (ISSN 1879-033X, PMID 21684150, DOI 10.1016/j.sbi.2011.05.003, lire en ligne, consulté le )
- Dasa Lipovsek et Andreas Plückthun, « In-vitro protein evolution by ribosome display and mRNA display », Journal of Immunological Methods, vol. 290, nos 1-2, , p. 51–67 (ISSN 0022-1759, PMID 15261571, DOI 10.1016/j.jim.2004.04.008, lire en ligne, consulté le )
- David T. F. Dryden, Andrew R. Thomson et John H. White, « How much of protein sequence space has been explored by life on Earth? », Journal of the Royal Society, Interface, vol. 5, no 25, , p. 953–956 (ISSN 1742-5689, PMID 18426772, PMCID PMC2459213, DOI 10.1098/rsif.2008.0085, lire en ligne, consulté le )
- O. Kuchner et F. H. Arnold, « Directed evolution of enzyme catalysts », Trends in Biotechnology, vol. 15, no 12, , p. 523–530 (ISSN 0167-7799, PMID 9418307, DOI 10.1016/S0167-7799(97)01138-4, lire en ligne, consulté le )
- S. Sen, V. Venkata Dasu et B. Mandal, « Developments in directed evolution for improving enzyme functions », Applied Biochemistry and Biotechnology, vol. 143, no 3, , p. 212–223 (ISSN 0273-2289, PMID 18057449, lire en ligne, consulté le )
- D. Dafydd Jones, « Triplet nucleotide removal at random positions in a target gene: the tolerance of TEM-1 beta-lactamase to an amino acid deletion », Nucleic Acids Research, vol. 33, no 9, , e80 (ISSN 1362-4962, PMID 15897323, PMCID PMC1129029, DOI 10.1093/nar/gni077, lire en ligne, consulté le )
- W. P. Stemmer, « Rapid evolution of a protein in vitro by DNA shuffling », Nature, vol. 370, no 6488, , p. 389–391 (ISSN 0028-0836, PMID 8047147, DOI 10.1038/370389a0, lire en ligne, consulté le )
- A. Crameri, S. A. Raillard, E. Bermudez et W. P. Stemmer, « DNA shuffling of a family of genes from diverse species accelerates directed evolution », Nature, vol. 391, no 6664, , p. 288–291 (ISSN 0028-0836, PMID 9440693, DOI 10.1038/34663, lire en ligne, consulté le )
- Manfred T. Reetz et José Daniel Carballeira, « Iterative saturation mutagenesis (ISM) for rapid directed evolution of functional enzymes », Nature Protocols, vol. 2, no 4, , p. 891–903 (ISSN 1750-2799, PMID 17446890, DOI 10.1038/nprot.2007.72, lire en ligne, consulté le )
- Daniel L. Hartl, « What can we learn from fitness landscapes? », Current Opinion in Microbiology, vol. 21, , p. 51–57 (ISSN 1879-0364, PMID 25444121, PMCID PMC4254422, DOI 10.1016/j.mib.2014.08.001, lire en ligne, consulté le )
- Ahmed H. Badran et David R. Liu, « In vivo continuous directed evolution », Current Opinion in Chemical Biology, vol. 24, , p. 1–10 (ISSN 1879-0402, PMID 25461718, PMCID PMC4308500, DOI 10.1016/j.cbpa.2014.09.040, lire en ligne, consulté le )
- Ashwani Kumar et Suren Singh, « Directed evolution: tailoring biocatalysts for industrial applications », Critical Reviews in Biotechnology, vol. 33, no 4, , p. 365–378 (ISSN 1549-7801, PMID 22985113, DOI 10.3109/07388551.2012.716810, lire en ligne, consulté le )
- William G. T. Willats, « Phage display: practicalities and prospects », Plant Molecular Biology, vol. 50, no 6, , p. 837–854 (ISSN 0167-4412, PMID 12516857, lire en ligne, consulté le )
- Hans Leemhuis, Viktor Stein, Andrew D. Griffiths et Florian Hollfelder, « New genotype-phenotype linkages for directed evolution of functional proteins », Current Opinion in Structural Biology, vol. 15, no 4, , p. 472–478 (ISSN 0959-440X, PMID 16043338, DOI 10.1016/j.sbi.2005.07.006, lire en ligne, consulté le )
- Kathryn D. Verhoeven, Olvia C. Altstadt et Sergey N. Savinov, « Intracellular detection and evolution of site-specific proteases using a genetic selection system », Applied Biochemistry and Biotechnology, vol. 166, no 5, , p. 1340–1354 (ISSN 1559-0291, PMID 22270548, DOI 10.1007/s12010-011-9522-6, lire en ligne, consulté le )
- Annalee W. Nguyen et Patrick S. Daugherty, « Evolutionary optimization of fluorescent proteins for intracellular FRET », Nature Biotechnology, vol. 23, no 3, , p. 355–360 (ISSN 1087-0156, PMID 15696158, DOI 10.1038/nbt1066, lire en ligne, consulté le )
- Yolanda Schaerli et Florian Hollfelder, « The potential of microfluidic water-in-oil droplets in experimental biology », Molecular bioSystems, vol. 5, no 12, , p. 1392–1404 (ISSN 1742-2051, PMID 20023716, DOI 10.1039/b907578j, lire en ligne, consulté le )
- Shannon A. Marshall, Greg A. Lazar, Arthur J. Chirino et John R. Desjarlais, « Rational design and engineering of therapeutic proteins », Drug Discovery Today, vol. 8, no 5, , p. 212–221 (ISSN 1359-6446, PMID 12634013, lire en ligne, consulté le )
- Corey J. Wilson, « Rational protein design: developing next-generation biological therapeutics and nanobiotechnological tools », Wiley Interdisciplinary Reviews. Nanomedicine and Nanobiotechnology, vol. 7, no 3, , p. 330–341 (ISSN 1939-0041, PMID 25348497, DOI 10.1002/wnan.1310, lire en ligne, consulté le )
- Lars Giger, Sami Caner, Richard Obexer et Peter Kast, « Evolution of a designed retro-aldolase leads to complete active site remodeling », Nature Chemical Biology, vol. 9, no 8, , p. 494–498 (ISSN 1552-4469, PMID 23748672, PMCID PMC3720730, DOI 10.1038/nchembio.1276, lire en ligne, consulté le )
- U. T. Bornscheuer et M. Pohl, « Improved biocatalysts by directed evolution and rational protein design », Current Opinion in Chemical Biology, vol. 5, no 2, , p. 137–143 (ISSN 1367-5931, PMID 11282339, lire en ligne, consulté le )
- Moshe Goldsmith et Dan S. Tawfik, « Directed enzyme evolution: beyond the low-hanging fruit », Current Opinion in Structural Biology, vol. 22, no 4, , p. 406–412 (ISSN 1879-033X, PMID 22579412, DOI 10.1016/j.sbi.2012.03.010, lire en ligne, consulté le )
- Mike M. Y. Chen, Christopher D. Snow, Christina L. Vizcarra et Stephen L. Mayo, « Comparison of random mutagenesis and semi-rational designed libraries for improved cytochrome P450 BM3-catalyzed hydroxylation of small alkanes », Protein engineering, design & selection: PEDS, vol. 25, no 4, , p. 171–178 (ISSN 1741-0134, PMID 22334757, DOI 10.1093/protein/gzs004, lire en ligne, consulté le )
- Carlos G. Acevedo-Rocha, Sabrina Hoebenreich et Manfred T. Reetz, « Iterative saturation mutagenesis: a powerful approach to engineer proteins by systematically simulating Darwinian evolution », Methods in Molecular Biology (Clifton, N.J.), vol. 1179, , p. 103–128 (ISSN 1940-6029, PMID 25055773, DOI 10.1007/978-1-4939-1053-3_7, lire en ligne, consulté le )
- Helge Jochens et Uwe T. Bornscheuer, « Natural diversity to guide focused directed evolution », Chembiochem: A European Journal of Chemical Biology, vol. 11, no 13, , p. 1861–1866 (ISSN 1439-7633, PMID 20680978, DOI 10.1002/cbic.201000284, lire en ligne, consulté le )
- Helge Jochens, Dirk Aerts et Uwe T. Bornscheuer, « Thermostabilization of an esterase by alignment-guided focussed directed evolution », Protein engineering, design & selection: PEDS, vol. 23, no 12, , p. 903–909 (ISSN 1741-0134, PMID 20947674, DOI 10.1093/protein/gzq071, lire en ligne, consulté le )
- Nicholas J. Turner, « Directed evolution drives the next generation of biocatalysts », Nature Chemical Biology, vol. 5, no 8, , p. 567–573 (ISSN 1552-4469, PMID 19620998, DOI 10.1038/nchembio.203, lire en ligne, consulté le )
- Philip A. Romero et Frances H. Arnold, « Exploring protein fitness landscapes by directed evolution », Nature Reviews. Molecular Cell Biology, vol. 10, no 12, , p. 866–876 (ISSN 1471-0080, PMID 19935669, PMCID PMC2997618, DOI 10.1038/nrm2805, lire en ligne, consulté le )
- Pietro Gatti-Lafranconi, Antonino Natalello, Sascha Rehm et Silvia Maria Doglia, « Evolution of stability in a cold-active enzyme elicits specificity relaxation and highlights substrate-related effects on temperature adaptation », Journal of Molecular Biology, vol. 395, no 1, , p. 155–166 (ISSN 1089-8638, PMID 19850050, DOI 10.1016/j.jmb.2009.10.026, lire en ligne, consulté le )
- H. Zhao et F. H. Arnold, « Directed evolution converts subtilisin E into a functional equivalent of thermitase », Protein Engineering, vol. 12, no 1, , p. 47–53 (ISSN 0269-2139, PMID 10065710, lire en ligne, consulté le )
- R. E. Hawkins, S. J. Russell et G. Winter, « Selection of phage antibodies by binding affinity. Mimicking affinity maturation », Journal of Molecular Biology, vol. 226, no 3, , p. 889–896 (ISSN 0022-2836, PMID 1507232, lire en ligne, consulté le )
- Fathima Aidha Shaikh et Stephen G. Withers, « Teaching old enzymes new tricks: engineering and evolution of glycosidases and glycosyl transferases for improved glycoside synthesis », Biochemistry and Cell Biology = Biochimie Et Biologie Cellulaire, vol. 86, no 2, , p. 169–177 (ISSN 0829-8211, PMID 18443630, DOI 10.1139/O07-149, lire en ligne, consulté le )
- Manoj Cheriyan, Matthew J. Walters, Brian D. Kang et Laura L. Anzaldi, « Directed evolution of a pyruvate aldolase to recognize a long chain acyl substrate », Bioorganic & Medicinal Chemistry, vol. 19, no 21, , p. 6447–6453 (ISSN 1464-3391, PMID 21944547, PMCID PMC3209416, DOI 10.1016/j.bmc.2011.08.056, lire en ligne, consulté le )
- G. MacBeath, P. Kast et D. Hilvert, « Redesigning enzyme topology by directed evolution », Science (New York, N.Y.), vol. 279, no 5358, , p. 1958–1961 (ISSN 0036-8075, PMID 9506949, lire en ligne, consulté le )
- Miguel D. Toscano, Kenneth J. Woycechowsky et Donald Hilvert, « Minimalist active-site redesign: teaching old enzymes new tricks », Angewandte Chemie (International Ed. in English), vol. 46, no 18, , p. 3212–3236 (ISSN 1433-7851, PMID 17450624, DOI 10.1002/anie.200604205, lire en ligne, consulté le )
- Svante Pääbo, Hendrik Poinar, David Serre et Viviane Jaenicke-Despres, « Genetic analyses from ancient DNA », Annual Review of Genetics, vol. 38, , p. 645–679 (ISSN 0066-4197, PMID 15568989, DOI 10.1146/annurev.genet.37.110801.143214, lire en ligne, consulté le )
- M. Höss, P. Jaruga, T. H. Zastawny et M. Dizdaroglu, « DNA damage and DNA sequence retrieval from ancient tissues », Nucleic Acids Research, vol. 24, no 7, , p. 1304–1307 (ISSN 0305-1048, PMID 8614634, lire en ligne, consulté le )
- Jesse D. Bloom et Frances H. Arnold, « In the light of directed evolution: pathways of adaptive protein evolution », Proceedings of the National Academy of Sciences of the United States of America, vol. 106 Suppl 1, , p. 9995–10000 (ISSN 1091-6490, PMID 19528653, PMCID PMC2702793, DOI 10.1073/pnas.0901522106, lire en ligne, consulté le )
- Alan M. Moses et Alan R. Davidson, « In vitro evolution goes deep », Proceedings of the National Academy of Sciences of the United States of America, vol. 108, no 20, , p. 8071–8072 (ISSN 1091-6490, PMID 21551096, PMCID PMC3100951, DOI 10.1073/pnas.1104843108, lire en ligne, consulté le )
- K. Salehi-Ashtiani et J. W. Szostak, « In vitro evolution suggests multiple origins for the hammerhead ribozyme », Nature, vol. 414, no 6859, , p. 82–84 (ISSN 0028-0836, PMID 11689947, DOI 10.1038/35102081, lire en ligne, consulté le )
- M. Sumper et R. Luce, « Evidence for de novo production of self-replicating and environmentally adapted RNA structures by bacteriophage Qbeta replicase », Proceedings of the National Academy of Sciences of the United States of America, vol. 72, no 1, , p. 162–166 (ISSN 0027-8424, PMID 1054493, lire en ligne, consulté le )
- D. R. Mills, R. L. Peterson et S. Spiegelman, « An extracellular Darwinian experiment with a self-duplicating nucleic acid molecule », Proceedings of the National Academy of Sciences of the United States of America, vol. 58, no 1, , p. 217–224 (ISSN 0027-8424, PMID 5231602, lire en ligne, consulté le )
- J. H. Marden, M. R. Wolf et K. E. Weber, « Aerial performance of Drosophila melanogaster from populations selected for upwind flight ability », The Journal of Experimental Biology, vol. 200, no Pt 21, , p. 2747–2755 (ISSN 0022-0949, PMID 9418031, lire en ligne, consulté le )
- William C. Ratcliff, R. Ford Denison, Mark Borrello et Michael Travisano, « Experimental evolution of multicellularity », Proceedings of the National Academy of Sciences of the United States of America, vol. 109, no 5, , p. 1595–1600 (ISSN 1091-6490, PMID 22307617, PMCID PMC3277146, DOI 10.1073/pnas.1115323109, lire en ligne, consulté le )
- Jeffrey E. Barrick, Dong Su Yu, Sung Ho Yoon et Haeyoung Jeong, « Genome evolution and adaptation in a long-term experiment with Escherichia coli », Nature, vol. 461, no 7268, , p. 1243–1247 (ISSN 1476-4687, PMID 19838166, DOI 10.1038/nature08480, lire en ligne, consulté le )
- Richard H. Heineman, Ian J. Molineux et James J. Bull, « Evolutionary robustness of an optimal phenotype: re-evolution of lysis in a bacteriophage deleted for its lysin gene », Journal of Molecular Evolution, vol. 61, no 2, , p. 181–191 (ISSN 0022-2844, PMID 16096681, DOI 10.1007/s00239-004-0304-4, lire en ligne, consulté le )
- Takuyo Aita, Norio Hamamatsu, Yukiko Nomiya et Hidefumi Uchiyama, « Surveying a local fitness landscape of a protein with epistatic sites for the study of directed evolution », Biopolymers, vol. 64, no 2, , p. 95–105 (ISSN 0006-3525, PMID 11979520, DOI 10.1002/bip.10126, lire en ligne, consulté le )
- Barrett Steinberg et Marc Ostermeier, « Environmental changes bridge evolutionary valleys », Science Advances, vol. 2, no 1, , e1500921 (ISSN 2375-2548, PMID 26844293, PMCID PMC4737206, DOI 10.1126/sciadv.1500921, lire en ligne, consulté le )
- F. H. Arnold, P. L. Wintrode, K. Miyazaki et A. Gershenson, « How enzymes adapt: lessons from directed evolution », Trends in Biochemical Sciences, vol. 26, no 2, , p. 100–106 (ISSN 0968-0004, PMID 11166567, lire en ligne, consulté le )
- Jesse D. Bloom, Alpan Raval et Claus O. Wilke, « Thermodynamics of neutral protein evolution », Genetics, vol. 175, no 1, , p. 255–266 (ISSN 0016-6731, PMID 17110496, PMCID PMC1775007, DOI 10.1534/genetics.106.061754, lire en ligne, consulté le )
Liens externes
- Groupes de recherche
- (en) Groupe de recherche de Dan Tawfik
- (en) Groupe de recherche d'Ulrich Schwaneberg
- (en) Groupe de recherche de Frances Arnold
- (en) Groupe de recherche de Huimin Zhao
- (en) Groupe de recherche de Darren Hart
- (en) Groupe de recherche de David Liu
- (en) Groupe de recherche de Douglas Clark
- (en) Groupe de recherche de Paul Balby
- (en) SeSaM-Biotech - Évolution Dirigée
- (en) Le Prof. Reetz explique le principe de l'Évolution Dirigée
- (en) Compagnie Codexis, Inc.