Recombinaison génétique
La recombinaison génétique est « le phénomène conduisant à l’apparition, dans une cellule ou dans un individu, de gènes ou de caractères héréditaires dans une association différente de celle observée chez les cellules ou individus parentaux »[1]. Cette définition recouvre deux processus complémentaires que sont les brassages intra et interchromosomiques, elle est toujours la définition utilisée en génétique, génétique des populations ou virologie, et peut être considérée comme synonyme de brassage génétique.
Cependant le développement de la biologie moléculaire a mené vers une interprétation différente de cette définition par une partie de la communauté scientifique. En biologie moléculaire le terme recombinaison génétique est souvent utilisé comme synonyme de la recombinaison de l'ADN, c'est-à-dire les processus par lesquels une molécule d'ADN (ou d'ARN) est coupée, puis jointe à une autre[2]. Les recombinaisons génétiques provoquées artificiellement sont également un outil essentiel en biologie moléculaire et génie génétique. En générant de nouvelles combinaisons génétiques, les recombinaisons naturelles sont un des mécanismes à l'origine de la diversité d'une population. Ce brassage génétique, facilité par la reproduction sexuée, est un des mécanismes essentiels de l'évolution des espèces.
Généralités
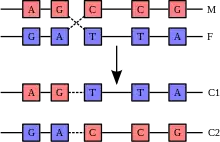
La recombinaison génétique est un échange d'information génétique entre deux génomes différents ou bien entre deux chromosomes. Il s'agit en général d'un échange entre fragments d'ADN. Mais chez certains virus comme celui de la grippe, il s'agit d'échange d'ARN. Cela permet de créer de nouvelles combinaisons génétiques donc des génomes nouveaux. La recombinaison est un phénomène naturel et universel dans le monde vivant, et c'est un des facteurs essentiels permettant de maintenir la diversité génétique dans une population. La formation de nouvelles combinaisons génétiques assure le brassage génétique et le maintien de la diversité génétique dans une population, ce qui augmente la possibilité pour une espèce de s'adapter à une modification de l'environnement. La recombinaison est donc l'un des processus essentiels de l'évolution des espèces.
- Chez les eucaryotes, elle se produit lors de la reproduction sexuée, grâce à la méiose (quand se forment les gamètes, et grâce à la fécondation) ou dans les cellules somatiques, lors de la mitose (recombinaison mitotique (en)).
- Chez les procaryotes (bactéries), elle se produit grâce à la conjugaison bactérienne, la transformation bactérienne, ou la transduction bactérienne.
- Chez les virus, la recombinaison peut avoir lieu au sein des cellules infectées par deux virus différents. L'apparition du nouveau virus H5N1 est par exemple, le résultat de recombinaisons génétiques.
Du fait du coût lié à la synthèse des enzymes nécessaires au processus, la maintenance de la recombinaison est nécessairement associée à un avantage évolutif. L'hypothèse la plus probable, du moins chez les eucaryotes, est liée à l'amélioration du brassage génétique au cours de la reproduction sexuée. En l'absence de recombinaison, les chromosomes seraient redistribués aléatoirement au cours de la méiose, mais les gènes liés sur le même chromosome resteraient toujours associés. La recombinaison, en permettant le réassortiment des haplotypes, permet de diminuer le déséquilibre de liaison et de créer de nouvelles combinaisons, augmentant ainsi la diversité génétique au sein de la population. L'évolution de la fonction de recombinaison se heurte néanmoins aux mêmes obstacles théoriques que l'évolution de la reproduction sexuée, et n'est susceptible de se produire que dans des conditions démographiques (taille de population) et génétiques (importance et directionalité des interactions génétiques) spécifiques, et pas encore clairement élucidées[3] - [4].
Chez les eucaryotes

Chez les espèces eucaryotes, le brassage génétique intervient au cours de la reproduction sexuée des espèces. Chaque individu possède deux allèles différents de chaque gène s'il est hétérozygote pour ces allèles (sinon on dit qu'il est homozygote). Lors d'une reproduction sexuée, le nouvel individu va hériter, pour chaque gène, d'un des deux allèles de son premier parent - sélectionné aléatoirement - et d'un des deux allèles de son autre parent. Le patrimoine génétique du nouvel individu est donc ainsi composé aléatoirement d'une partie du patrimoine de chacun de ses deux parents. Ceci permet au matériel génétique de se répandre au sein de l'espèce.
Il existe deux principaux types de brassage génétique, qui ont lieu lors de la méiose :
- Recombinaisons intra-chromosomiques : Les deux chromatides d'une même paire portent des allèles différents à un certain nombre de locus. Au cours de la prophase de 1re division méiotique, les chromosomes homologues s'apparient et s'enchevêtrent au niveau des chiasmas. Il se produit des échanges de segments entre ces chromosomes. Ce phénomène est l'enjambement (ou crossing-over) : un allèle d'un chromosome peut être échangé avec l'allèle porté par le chromosome homologue. Tous les gènes situés sur une paire de chromosomes peuvent être « brassés » grâce à l'enjambement ce qui modifie l'association d'allèles portés par chacun des chromosomes. Ce brassage entre allèles d'une paire homologue est qualifiée d'intrachromosomique. Un chromosome comporte en moyenne 1 000 gènes, alors il y aurait théoriquement 21000 possibilités. Mais ce calcul ne prend pas en compte les probabilités relatives au nombre de chevauchements possibles sur un même chromosome : il n'y en a en moyenne que de 1 à 5 par chromosome.
- Recombinaisons inter-chromosomiques : Elles ont lieu pendant la méiose, en anaphase ou lorsque la cellule se sépare en deux, il y a une séparation aléatoire des chromosomes: si par exemple il y a deux paires de chromosomes noté A et B et 1 et 2 il existe 4 possibilités soit A avec 1, B et 1, B et 2 ou A et 2. Chez l'homme et la femme il y a 23 paires de chromosomes, ce qui fait 223 possibilités soit plus de 8 millions. La dernière étape du brassage génétique est la fécondation: la rencontre au hasard des gamètes. Elle multiplie encore plus les possibilités, soit dans notre exemple 8 millions × 8 millions = 64 000 milliards.
Donc le brassage interchromosomique et la fécondation apportent plus de 64 000 milliards (64 × 1012) combinaisons génétiques originales et donc de descendants différents si on exclut les cas de forte consanguinité. En rajoutant les recombinaisons intra-chromosomiques, le nombre de combinaisons originales et donc de descendants différents augmente encore et rend la probabilité d'avoir deux individus avec le même génome (hors vrais-jumeaux) quasi nulle.
Chez les bactéries
Il existe trois mécanismes possibles : la conjugaison bactérienne, la transformation des bactéries et la transduction. Grete Kellenberger a fait des découvertes majeures dans ce domaine en étudiant le phage lambda: elle met en évidence la récombinaison est due à un échange physique d'ADN, plutôt qu'à une réplication selective[5].
Chez les virus
L'apparition de nouveau virus (H5N1) est aussi un exemple de recombinaison génétique.
Intérêts économique et médical
Les recombinaisons génétiques ont donc pour effet de créer des variétés nouvelles d'animaux ou de végétaux pouvant être intéressantes en agriculture. Ces variétés recombinées peuvent être obtenus par croisement naturel, croisement artificiel. On peut aussi provoquer des recombinaisons génétiques par le biais de technologies de biologie moléculaire et de manipulation de l'ADN. On parle alors d'un OGM.
Niveau moléculaire
Celle-ci nécessite la présence de séquences homologues entre deux régions de l'ADN.
Le complexe RecBCD déroule l'ADN et coupe un brin à proximité d'un site chi. Une extrémité 5' simple brin est produite et va envahir la molécule homologue. La protéine RecA va alors se charger de rechercher une séquence homologue : elle se fixe sur le simple brin avec SSB et réunit les deux séquences. La formation d'un hétéroduplex se termine par l'action de la ligase. Il se produit ensuite une rotation de l'hétéroduplex conduisant à la formation de la jonction de Holliday. Sous l'action de la protéine RuvAB, cette jonction va migrer et va être coupée (résolution) grâce à RuvC. Il y a deux coupures possibles conduisant soit à un enjambement (recombinaison entre allèles), soit à une zone hétéroduplex (la probabilité de survenue est identique).
Complexe Rad51 : rôle de la protéine RecA.
Recombinaison homologue
Le terme recombinaison homologue, correspond à un évènement de recombinaison génétique entre deux séquences identiques situées sur 2 molécules d'ADN différentes, ou distantes l'une de l'autre sur la même molécule. Ce processus est très fréquent chez la levure et donc largement utilisé comme outil de biologie moléculaire dans ce cas.
Recombinaison illégitime
La recombinaison illégitime (ou non homologue) est un processus cellulaire conduisant à la réunion de deux fragments d'ADN ne présentant pas ou très peu d'homologie de séquence. Elle est à l'origine de diverses anomalies du génome (délétion, insertion, translocation, amplification génique, intégration virale) impliquées dans l'apparition de maladies génétiques ou le développement tumoral. Elle permet l'élimination des cassures double-brin accidentelles de la chaîne nucléotidique, consécutives à des erreurs du métabolisme normal de l'ADN (réplication, transcription, réparation) ou à la présence de lésions sur l'ADN produites par les agents génotoxiques
Recombinaison spécifique d’un site
Étape de reconnaissance guidé par une enzyme de restriction. L'appariement entre les deux molécules en cours de recombinaison n'est pas épigé. L'enzyme de restriction reconnaît des séquences nucléotidiques spécifiques.
Notes et références
- « http://www.culture.gouv.fr/culture/dglf/terminologie/repertoireJO220900/accueil.htm »(Archive.org • Wikiwix • Archive.is • Google • Que faire ?) définition historique, et retenue par le Journal officiel de la République française: Journal officiel de la République française du 22 septembre 2000, Répertoire terminologique 2000]
- Biologie cellulaire et moléculaire, de Gerald Karp, 2007, publié par De Boeck Université, (ISBN 2804145379), 9782804145378
- de Visser JA, Elena SF. The evolution of sex: empirical insights into the roles of epistasis and drift. Nat Rev Genet. 2007 139-149.
- Agrawal AF. Evolution of sex: why do organisms shuffle their genotypes? Curr Biol. 2006 R696-704.
- (en) Kellenberger G., Zichichi, M.L., Weigle,, « Exchange of DNA in the recombination of bacteriophage lambda » », Proc. Natl. Acad. Sci. USA, no no 47, , p. 869-878