Virus d'Epstein-Barr
Le virus d'Epstein-Barr (aussi appelé EBV) ou virus de l’herpès 4 (HHV-4) est un virus de la famille des Herpesviridae. Il fait partie de la sous-famille des Gammaherpesvirinae (en). Le virus d'Epstein-Barr cause plusieurs maladies dont la mononucléose infectieuse et le lymphome de Burkitt.
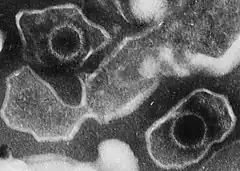
entourée par la membrane plasmique.
| Type | Virus |
|---|---|
| Groupe | Groupe I |
| Famille | Herpesviridae |
| Sous-famille | Gammaherpesvirinae |
| Genre | Lymphocryptovirus |
Le virus cible les lymphocytes B humains via leur récepteur CD21 (en). La protéine EBNA-2 (en) qu'il produit lors de l'infection de ces cellules, peut recruter des facteurs de transcription (FT) humains et ainsi causer des maladies auto-immunes comme le lupus ou le diabète de type 1[1] mais aussi la sclérose en plaques[2] - [3] - [4].
Le laboratoire Moderna a engagé les essais d'un candidat vaccin à ARNm contre ce virus[2].
Histoire
Le virus a été découvert en 1964 par l’analyse d’une biopsie d’un spécimen de tumeur en microscopie électronique, il fut nommé ainsi en l’honneur de Michael A. Epstein et de son étudiante Yvonne Barr. En 1968, le Dr Henle et son équipe de recherche établirent un lien entre la mononucléose infectieuse et le virus d'Epstein-Barr. Ils observèrent qu’une technicienne du laboratoire présentait une mononucléose infectieuse ainsi qu’une séroconversion vis-à-vis du virus.
Réservoir, tropisme et transmission
Le réservoir du virus d'Epstein-Barr est strictement humain. Le virus est transmis par la salive. Le virus d'Epstein-Barr se multiplie dans les cellules de l’oropharynx. Quoique les cellules de l’oropharynx soient permissives à la réplication du virus, des données récentes suggèrent que les lymphocytes B retrouvés dans l’oropharynx seraient le site d’infection primaire. Les lymphocytes B sont donc les cellules cibles du virus d'Epstein-Barr, et plus précisément via leur antigène de surface CD21 (en), véritable porte d'entrée du virus.
Épidémiologie et maladies
Le virus d'Epstein-Barr fait partie des virus humains les plus communs et est retrouvé partout dans le monde. Le virus infecte 80-90 % des adultes dans le monde[5]. Aux États-Unis, 95 % des adultes entre 35 et 40 ans ont été infectés.
Le virus d'Epstein-Barr est plus fréquemment acquis pendant l’enfance dans les pays sous-développés (> 90 % des enfants d’âge préscolaire). Aux États-Unis et dans les autres pays développés, beaucoup de personnes ne sont pas infectées lors de l’enfance, mais le sont plutôt lors de l’adolescence ou durant la vie adulte. Le virus cause plusieurs maladies à travers le monde.
Les porteurs du virus sont contagieux par la salive, et des études tendent à prouver que le virus est transmissible par les relations sexuelles[6].
Le risque de sclérose en plaques est multiplié par 32 après une infection par le virus d'Epstein-Barr[7].
| Personne immunocompétente (saine) | Personne immunodéprimée |
|---|---|
| Mononucléose infectieuse | Syndrome de Purtilo |
| Lymphome de Burkitt | Leucoplasie chevelue de la langue |
| Carcinome nasopharyngé | Lymphome de Burkitt |
| Lymphome T centro-facial | Lymphome immunoblastique |
| Maladie de Hodgkin | Maladie de Hodgkin |
Cependant, leur prédominance diffère selon la région du monde où ils sont retrouvés. Par exemple, pour le lymphome de Burkitt, le virus existe à l’état hyperendémique dans les régions hautement atteintes de paludisme comme l’Afrique tropicale.
Le virus d'Epstein-Barr cause la mononucléose infectieuse. Cependant, la maladie est rarement retrouvée dans les pays sous-développés. Aux États-Unis, lorsque l’infection par le virus a lieu durant l’adolescence ou à l’âge adulte, 35 % à 50 % des cas développent la mononucléose infectieuse. EBV a un rôle pathogène important dans le lymphome de Burkitt qui est retrouvé de façon endémique en Afrique. Le virus est impliqué dans 97 % des cas de lymphome de Burkitt en Afrique et en Papouasie-Nouvelle-Guinée dans les zones où les cas de paludismes sont fréquents[5].
De plus, deux types de virus sont observés soit EBV-1 et EBV-2 (ou bien type A et type B) et sont retrouvés partout dans le monde. Les études séroépidémiologiques et l’isolation du virus suggèrent que la plupart des personnes en Europe et Amérique du Nord sont infectées par le virus de type 1⁵ et le type B est principalement restreint à l’Afrique mais tous les deux sont répandus. Les maladies causées sont les mêmes. Cependant les gènes exprimés lors de la latence présentent quelques différences.
Certains cas d'allergies médicamenteuses sont également provoqués par l'EBV[5].
Diagnostic
Il existe des méthodes de diagnostic sérologique qui se basent sur la présence d'anticorps dirigés contre certaines protéines du virus ainsi que des méthodes moléculaires. Les méthodes sérologiques utilisent des tests tels la méthode ELISA ou bien la détection par immunofluorescence. Pour les méthodes moléculaires, des tests tels que le PCR, l’analyse de type Southern blot ou le NASBA sont utilisés.
Génome et structure
Le génome du virus d'Epstein-Barr est un ADN linéaire double brin de 172 kilopaires de base. Le génome encode approximativement 100 gènes dont 10 exprimés lors de la phase de latence du virus. Le virus d'Epstein-Barr consiste en un cœur protéique de forme toroïdal enveloppé avec l’ADN double brin. De plus, le virus possède une nucléocapside icosaédrique contenant 162 capsomères, une protéine d’enveloppe amorphe entourant la capside et une protéine d’enveloppe externe constituée de façon prédominante par une glycoprotéine unique : gp350/220.
Protéines virales
Le virus va exprimer lors de sa réplication des antigènes précoces (LYDMA, EBNA et EMA), lors de la duplication du matériel (EA) et des antigènes tardifs (LMA et VCA).
- LYDMA (Lymphocyte-defined membran antigen) est un antigène présent à la surface de la cellule infectée qui est la cible des lymphocytes T cytotoxiques.
- EBNA (Epstein-Barr nuclear antigen) est un antigène nucléaire.
- EMA (Early membran antigen) est un antigène précoce de la membrane cellulaire.
- EA (Early antigen) correspond à des protéines enzymatiques virales qui assurent la réplication du génome.
- LMA (Late membran antigen) est un antigène tardif de la membrane.
- VCA (Virus capsid antigen) est la protéine de la capside virale.
Certains gènes seront exprimés durant la latence (EBNA-1, EBNA-2, EBNA-3 EBNA-LP, LMP-1, LMP-2, EBER-1 et EBER-2)
- EBNA-1 est requise pour la réplication épisomale et la maintenance du génome viral
- EBNA-2 est essentiel au processus d’immortalisation des lymphocytes B et à l’expression des protéines EBNA- 1 et EBNA-3. La variation dans les protéines EBNA-2 cause les variations biologiques les plus significatives entre deux types de virus d'Epstein-Barr. EBNA-2 active en trans l’expression du marqueur d’activation CD23 du lymphocyte B et régule positivement l’expression du récepteur CD21 du virus, c-fgr et les gènes de latence LMP-1 et LMP-2.
- EBNA-3 est très polymorphique et diffère selon le type de virus. Il est constitué d’une famille de 3 gènes de haut poids moléculaire localisée en tandem sur le génome du virus d'Epstein-Barr.
- EBNA-LP est un ensemble de protéines extrêmement polymorphiques. Cependant la fonction de EBNA-LP demeure floue, elle pourrait jouer un rôle dans le traitement de l'ARN ou être associée avec certaines protéines de régulation nucléaire.
- LMP-1 est exprimé en absence de EBNA-2 pendant l’activation du cycle lytique des lymphocytes B. Cette protéine protège les lymphocytes B infectés de l’apoptose en partie par l’activation de l’oncogène bcl-2. De plus, l’action de transformation de LMP-1 semble impliquer les protéines de signalisation de TRAF (tumor necrosis factor receptor-associated factors)
- LMP-2 est une protéine de membrane contenant 12 domaines transmembranaires hydrophobiques qui est colocalisée avec LMP-1 dans la membrane plasmique des lymphocytes infectés.
- EBER-1 et EBER-2 sont les ARN du virus d'Epstein-Barr les plus abondants durant la latence des cellules B infectées. Ces petits ARN sont codés sous forme non polyadénylé. La majorité des EBER sont localisés au noyau et où ils sont complexés avec la protéine La. (La protéine La est une protéine ubiquitaire dans les cellules eucaryotes, et est associé à l’extrémité 3’ terminal des nouveaux ARN synthétisés.)
De plus, il existe aussi des glycoprotéines associées à la membrane cellulaire ainsi qu’à l’infectivité. La glycoprotéine gp340/220 en est la glycoprotéine majeure de ce groupe et est aussi associée à l’induction d’anticorps neutralisants.
Réplication et latence
La pénétration du virus à l’intérieur des lymphocytes B se fait par interaction entre la glycoprotéine gp 350/220 sur le virus et la protéine de surface CD21 chez le lymphocyte. L’adsorption du virus à la surface des lymphocytes est suivie par endocytose du virus d'Epstein-Barr. Une seconde glycoprotéine gp85 médie la fusion du virus avec la membrane cellulaire ce qui cause la relâche de la nucléocapside à l’intérieur du cytoplasme du lymphocyte B. Une troisième glycoprotéine gp42 est essentielle pour la pénétration des lymphocytes B par son interaction avec les molécules du CMH (complexe majeur d'histocompatibilité) de classe II à la surface de la cellule.
Le déshabillage de la nucléocapside et le transport du génome au noyau de la cellule sont moins bien compris. Le génome du virus d'Epstein-Barr est répliqué avec l’aide de l’ADN polymérase cellulaire durant la phase S du cycle cellulaire. Par la suite, le génome subsiste sous forme d’épisome.
La majorité des lymphocytes B en phase de latence n’entrent pas en cycle lytique cependant, ils peuvent être induits à y entrer soit par des esters de phorbol, des ionophores de calcium ou encore par des immunoglobulines membranaires à la surface de la cellule. Les gènes qui sont transcrits hâtivement, sont transcrits en absence de nouvelles protéines virales dans la cellule infectée. EBV exprime durant les étapes tardives du cycle lytique les gènes qui codent les protéines de structure.
La latence est caractérisée par trois processus distincts : la persistance virale, l’expression limitée du virus altérant sa prolifération et la conservation du potentiel de réactivation pour la réplication lytique. L’épisome est répliqué de façon semi conservative. Lors de la latence 10 gènes sont exprimés soit 6 EBNA, 2 LMP et 2 EBER afin d’assurer le maintien de l’état d’immortalité.
Bibliographie
- Richman D. D., Whitley J. R., Hayden G. F., 2002, Clinical Virology (second edition), ASM Press, pp. 479-486
- Hervé J. A. Fleury,2002, Virologie humaine (4e édition), Paris: Masson, pp.103-104
- Collier, L.H. et J.S. Oxford, 2006, Human virology: a text for students of medicine, dentistry, and microbiology (third edition), Oxford University Press, pp.153-157
- Specter, S. et al., 2009, Clinical Virology Manual (fourth edition), ASM Press, pp. 209-210
- Mammette, A., 2002, Virologie médicale, Presses universitaires de Lyon, pp.523-534
- Evans, S.A. et R. Kaslow, 1997, Viral infections of humans : epidemiology and control(fouth edition), Plenum Medical Book, pp.255-258
- http://www.phac-aspc.gc.ca/msds-ftss/msds62f-fra.php, consulté le
- https://www.cdc.gov/ncidod/diseases/ebv.htm, consulté le
- http://emedicine.medscape.com/article/963894-overview, consulté le
- Tsuchiya S., Diagnosis of Epstein-Barr virus-associated diseases, Critical Review in Oncology Hematology (vol.44), 2002,pp.227-238
Références
- Fabienne Rigal, « Le virus Epstein-Barr lié à 7 maladies auto-immunes dont le lupus et le diabète », sur Le Quotidien du médecin, (consulté le ).
- Claire Manière, « Sclérose en plaques : le virus d’Epstein-Barr en serait la cause principale », sur Trust my science, (consulté le ).
- Ouest-France avec AFP, « Sclérose en plaques. Un virus pourrait être à l’origine de la maladie, selon des chercheurs », sur ouest-france.fr, Ouest-France, (consulté le )
- (en) William H. Robinson et Lawrence Steinman, « Epstein-Barr virus and multiple sclerosis », Science, (DOI 10.1126/science.abm7930, lire en ligne
 , consulté le ).
, consulté le ). - Stéphane Biacchesi, Christophe Chevalier, Marie Galloux, Christelle Langevin, Ronan Le Goffic et Michel Brémont, Les virus : Ennemis ou alliés ?, Versailles, Quæ, coll. « Enjeux Sciences », , 112 p. (ISBN 978-2-7592-2627-6, lire en ligne), III. Quels sont les modes de transmission des virus ?, chap. 8 (« La latence virale »), p. 64-65, accès libre.
- (en) « Sexual History and Epstein-Barr Virus Infection », (consulté le )
- (en) Kjetil Bjornevik, Marianna Cortese, Brian C. Healy, Jens Kuhle, Michael J. Mina et al., « Longitudinal analysis reveals high prevalence of Epstein-Barr virus associated with multiple sclerosis », Science, vol. 375, no 6578, , p. 296-301 (DOI 10.1126/science.abj8222).