Remodelage du patron de développement
En génétique, le remodelage du patron de développement est lorsque l' altération d'un patron de développement originel aboutit à l'apparition d'un nouveau phénotype avec un patron de développement altéré. Bien que les mécanismes qu'il englobe soient connus depuis longtemps, le terme « remodelage du patron de développement » (de l'anglais developmental repatterning) n'a été introduit qu'en 2011 par le zoologiste Wallace Arthur afin de trouver un terme générique mettant en relation tous les grands concepts de la Biologie évolutive du développement (Evo-Devo). En considérant la formation d'un individu comme le résultat d'un patron de développement (developmental patterning), il en a alors déduit que la modification de ce patron à l'échelle évolutionnaire pouvait être vu comme un remodelage (developmental repatterning) causé par des facteurs à la fois génétiques et/ou environnementaux[1] - [2].
On reconnait aujourd'hui quatre formes de remodelage, l'hétérochronie, l'hétérotopie, l'hétérométrie et l'hétérotypie qui peuvent toutes être à la fois observables au niveau moléculaire comme au niveau morphologique.
Causes du Remodelages
La mutation
La mutation est sans doute la cause principale du remodelage. Définie comme l'ensemble des processus aboutissant à une modification de la séquence d'ADN d'un organisme, son impact dans l'évolution et tout particulièrement dans le remodelage peut être totalement différent selon sa taille (mutation ponctuelle, de centaines de paires de base ou de millier de paires de base), son type et surtout sa localisation. Pour entraîner une modification dans le patron de développement, elle doit tout d'abord affecter un gène de développement. Pour avoir un impact dans l'évolution, elle doit être héréditaire, donc être portée sur les cellules germinales, et doit également affecter la fitness de l'organisme porteur de cette mutation.
Il est néanmoins important de noter que la taille de la mutation n'implique pas automatiquement un impact d'une même envergure équivalente sur le développement. Selon sa localisation, la mutation d'une seule paire de base peut avoir un effet bien plus drastique sur le développement que la mutation de centaines de paires de bases.
Les actions principales d'une mutation sur le remodelage du patron du développement se font lorsque la mutation a lieu dans une région régulatrice ou dans une région codante d'un gène. Si elle apparaît ailleurs dans le génome, l'effet de la mutation sera le plus souvent neutre, sauf si elle influence indirectement la régulation génique. En affectant la région régulatrice d'un gène, la mutation peut changer la capacité de liaison du ou des facteurs de transcription et donc changer l'expression du gène que ce soit au niveau de sa fréquence, de sa quantité ou son moment d'expression. Un gène peut comprendre plusieurs zones régulatrices selon ses différents rôles dans le développement, une mutation sur une de ces régions peut ainsi modifier l'expression d'un seul des rôles du gène dans le développement.
En affectant la zone codante d'un gène, la mutation agit directement sur la ou les protéines issues de son expression et impacte donc la fonction même du gène. Ce genre de mutation entraîne une modification de l'ARNm transcrit et parfois la traduction d'une protéine différente (selon la redondance du code génétique) ou tronquée (en cas de modification du cadre de lecture)[3] - [2].
La méthylation
La méthylation de l'ADN correspond à l'ajout réversible d'un groupe méthyle sur quelques acides nucléiques, sans pour autant changer la séquence nucléotidique. La méthylation intervient dans la régulation de l'expression génique en inhibant ou en favorisant la transcription d'un gène. Un patron de méthylation est propre à chaque individu et est héréditaire. Chez les mammifères, il peut également être influencé par les facteurs environnementaux[3].
La plasticité phénotypique

Longtemps considérée comme d'influence mineure voire nulle, la plasticité phénotypique est aujourd'hui reconnue comme très importante dans le développement et dans la modification du patron de développement. La plasticité phénotypique peut être définie comme la capacité d'un organisme à répondre à un signal environnemental. Quand le changement s'observe lors du développement d'un embryon ou d'une larve, on peut également parler de plasticité développementale. Ainsi, pour un même individu et donc un même génotype, différents phénotypes sont possibles selon l'impact de l'environnement sur le patron de développement. Bien que pour que cette plasticité soit héréditaire et joue dans l'évolution il faut une assimilation génétique, le rôle de l'environnement est alors tout de même considéré comme plus important que la génétique dans le remodelage. Sur le long terme, tous les événements environnementaux pouvant influencer la capacité d'un organisme à être plastique sont des causes de remodelage[1] - [3].
Les Types de Remodelage
L'hétérochronie
L'hétérochronie est l'un des mécanismes les plus connus et qui a pendant longtemps été considéré, à tort, comme une des causes principales du remodelage. L'hétérochronie concerne toutes modifications d'ordre chronologique qui peuvent apparaître lors du développement. Par modification chronologique on entend l'apparition plus précoce ou au contraire plus tardive d'un événement ou bien un changement dans la durée d'un processus développemental.

L'hétérochronie peut être locale ou globale et existe à tous les niveaux, que ce soit au niveau moléculaire en affectant par exemple l'expression d'une protéine ou morphologique comme pour la croissance d'un organe. Elle est également observable à toutes les phases du développement, il est néanmoins plus fréquent de la retrouver dans les stades les plus avancés[2] - [4] - [5].
Bien qu'il existe plusieurs types d'hétérochronie, tels que la progénèse ou l'hypermorphose, l'un des plus connus est la néoténie qui correspond à un retard du développement somatique par rapport au système reproducteur. Il s'agit donc d'une hétérochronie globale qui touche l'ensemble de l'organisme. Chez la plupart des amphibiens, la métamorphose marque le passage de l'état larvaire à l'état adulte et est majoritairement due à la réponse des tissus face à une augmentation de thyroxine sécrétée par la thyroïde. Le phénomène de néoténie apparaît lorsque les tissus ne répondent plus à cette sécrétion de thyroxine, le développement somatique est alors retardé, voire annulé, mais la maturation sexuelle, indépendante à la thyroxine, reste inchangée. Ce retard peut être permanent ou bien facultatif comme pour le cas de axolotl. Chez l'axolotl, le niveau de thyroxine sécrété est insuffisant, il reste donc au stade larvaire toute sa vie mais est capable de se reproduire. Néanmoins, en cas d'exposition artificielle à de la thyroxine, la larve d'axolotl peut alors se métamorphoser et devenir une salamandre terrestre[2] - [6].
L'hétérotopie

L'hétérotopie est un autre célèbre type de remodelage qui concerne cette fois tout changement d'arrangement spatial observable lors du développement. En d'autres termes, on parle d'hétérotopie lorsqu'on observe une apparition, une disparition ou tout simplement un changement dans la localisation d'un processus développemental ou dans la formation d'un organe[3] - [2].
L'apparition de la carapace de tortue est un exemple d'hétérotopie. Lors du développement, les cellules à l'origine de la formation des côtes migrent latéralement par rapport aux somites et non ventralement pour former une cage thoracique, comme c'est le cas chez les autres vertébrés. Cette migration est permise grâce à la crête de la carapace, qui est une excroissance présente au-dessus des bourgeons de membre, qui va attirer les cellules dorsalement et va ensuite permettre aux côtes de fusionner entre elles, ainsi qu' au derme osseux, et donc de participer à la formation de la carapace[4]. Le développement latéral des côtes permet également l'internalisation des scapulas dans la cage thoracique alors qu'elles sont externes chez les autres vertébrés[2].
L'hétérométrie
L'hétérométrie concerne tout type de remodelage qui affecte le patron du développement d'un point de vue quantitatif, c'est-à-dire par la taille, la concentration ou le nombre. À l'instar des autres concepts du remodelage, l'hétérométrie ne peut s'observer que d'un point de vue relatif, en comparant deux lignées ou bien entre différentes parties de l'organisme.
Dans l'histoire de l'évolution, un phénomène d'hétérométrie peut être unidirectionnel ou bien bidirectionnel. L'évolution de la taille du cerveau entre l'australopithèque et l’homo-sapiens est un exemple d'hétérométrie unidirectionnelle car, d'après nos connaissances, le cerveau n'a fait qu'augmenter en taille (hétérométrie positive) pour arriver à la taille actuelle du cerveau humain[2]. Néanmoins, il est bien plus fréquent de retrouver des évènements d'hétérométries bidirectionnelles, c'est-à-dire à la fois positives et négatives, dans l'évolution. Il arrive également que l’apparition d'un caractère soit accompagné de la perte d'un autre, on parle alors d'hétérométrie compensatoire.
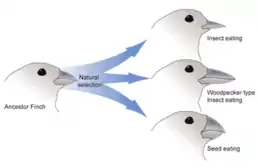
Les différences morphologiques du bec chez les pinsons des îles Galapagos (Pinsons de Darwin) est un exemple d'hétérométrie souvent utilisé. En fonction du mode de vie et de l'environnement où évolue l'animal, son bec adopte une forme et une taille différente. Selon des études récentes effectuées par Abzhanov et ses collègues, l'une des causes principales de ces différences morphologiques réside dans les changements de patron d'expression des protéines Bone morphogenetic protein 4 (BMP4) et calmoduline, impliquées entre autres dans la caractérisation du squelette facial et du bec[7] - [8]. Les changements peuvent être hétérométriques, c'est-à-dire dans la concentration des protéines, mais également hétérochroniques et hétérotopiques. Une expression de BMP4 précoce et en forte concentration induit un bec petit et large, adapté pour se nourrir de graines tandis qu'une expression retardée et plus faible de BMP4 induit un bec long et fin adapté à la recherche d'insecte et de fruits dans les fleurs de cactus[6]. Des résultats similaires ont été trouvés pour l'expression de la calmoduline.
L'hétérotypie
Le concept d'hétéroptypie est un concept relativement nouveau en Evo-Dévo et un peu différent, bien que totalement complémentaire, avec les fonctionnements des trois autres genres de remodelage. En effet, alors que l'hétérochronie, l'hétérométrie et l'hétérotypie agissent en modifiant la région régulatrice d'un gène, l'hétérotypie agit directement sur la région codante du gène ce qui induit une modification dans les propriétés fonctionnelles de la protéine synthétisée. La protéine produite pouvant être un facteur de transcription, une hétérotypie peut donc également impliquer indirectement un changement dans la régulation génique. L'hétérotypie consiste donc en l'apparition d'une nouvelle fonction ou d'un nouveau type, sans nécessairement impliquer une modification chronologique, spatiale ou quantitative du développement. Chez les Cnidaire, l'apparition d'un nouveau type cellulaire, la cellule urticante (ou nématocyste), est propre à leur embranchement et est un exemple d'hétérotypie[2].
Cause plausible de l'apparition de la grossesse utérine des mammifères
L'une des principales caractéristiques des femelles chez les mammifères est leur aptitude à allaiter leur progéniture ainsi que la gestation intra-utérine du fœtus. L'apparition de cette capacité serait en partie due à l'acquisition de nouvelles fonctions chez la prolactine ainsi qu'à sa synthèse dans les cellules utérines. La prolactine joue un rôle crucial pour la gestation. Que ce soit en induisant la différenciation des cellules épithéliales de l'utérus ou en régulant la réaction immunitaire et inflammatoire (permettant ainsi au fœtus de ne pas être considéré comme un corps étranger), la prolactine agit à différents stades dans le développement et la dans la vie d'une femelle pour rendre son utérus apte à accueillir un fœtus. L'acquisition de cette fonction durant l'évolution se serait faite grâce à la mutation du gène homéotique Hoxa11 des mammifères et à son expression dans les cellules destinées à former le placenta.
La mutation du gène Hoxa11 aurait permis son association avec le facteur de transcription Foxo1 a engendrant ainsi une régulation positive de la prolactine dans les cellules épithéliales de l'utérus. En effectuant une stratégie de perte de fonction chez la souris, il a été démontré qu'en l'absence d'expression du gène Hoxa11, la prolactine n'était pas synthétisée dans les cellules utérines. Cette expérience a ainsi permis la confirmation de l'hypothèse selon laquelle l'une des nouveautés évolutives les plus importantes aboutissant à l'apparition des mammifères est la modification hétérotypique de la séquence d'ADN du gène Hoxa11[3].
Cause de l'appartenance des insectes aux hexapodes

Dans la plupart des embranchements appartenant à la famille des arthropodes, les individus présentent au moins quatre paires de pattes alors que les insectes n'en présentent que trois, une sur chaque segment thoracique. Cette particularité serait due à une mutation dans la séquence du gène Ultrabithorax (Ubx), gène impliqué dans l'identification du troisième segment thoraciques ainsi que dans les segments abdominaux, entraînant l'apparition d'une région polyalanine dans la protéine Ubx. Alors qu'à l'origine le gène Ubx n'influence pas l'expression du gène distal-less (impliqué dans la formation d'appendices dans les segments), l'altération de la protéine entraîne une répression du gène distal-less dans les segments de l'abdomen et donc l'absence de pattes ou autres appendices dans cette région.
L'altération hétérotypique du gène Ubx a donc apporté une nouvelle fonction à la protéine synthétisée expliquant ainsi pourquoi les insectes n'ont que six pattes[3].
La Complexité du remodelage
La combinaison de plusieurs types
Au sein d'un même organisme, le remodelage du patron du développement peut être observé à différents niveaux, que ce soit au niveau moléculaire pour l'expression génique ou l'action d'une protéine, au niveau cellulaire ou bien au niveau de l'organisme entier en cas de changement morphologique. De ce fait, à l'échelle individuelle, une simple modification du développement peut être un type de remodelage différent selon le niveau étudié[2]. C'est le cas dans l'exemple étudié dans la section précédente sur la mutation hétérotypique du gène Ubx. En plus d'apporter une nouvelle fonction au gène Ubx, cette mutation entraîne la disparition des appendices au niveau de l'abdomen. D'un point de vue morphologique, c'est donc également un cas d'hétérométrie et d'hétérotopie.
De même, il peut arriver qu'à un niveau simple d'organisation on observe plusieurs types de changements pour un même événement. Par exemple une altération morphologique hétérométrique impliquant l'augmentation de la taille d'un organe va souvent indirectement impliquer un certain degré d'hétérotopie et d'hétérochronie car l'organe en question prendre plus de place et aura sans doute un délai de formation plus long.
Ainsi, à l'échelle de l'Evolution, un remodelage peut se révéler parfois très complexe. Bien que chaque type de remodelage peut être à lui seul à l'origine d'une nouveauté évolutive, sur le long terme, il n'est que rarement la résultante d'un seul de ces mécanismes. Le plus souvent, c'est un processus évolutif composé, dans lequel on peut observer une combinaison entre différents évènement d'hétérochronie, d'hétérotypie, d'hétérométrie ou d'hétérotopie[2].
Plusieurs remodelages, un seul résultat
Il arrive également que différents types de remodelage, agissant indépendamment les uns des autres, aboutissent à un même résultat.
Chez les vertébrés, la signalisation Sonic Hedgehog (shh) a un rôle crucial durant différents stades du développement, que ce soit par exemple lors de la myogénèse ou lors de la neurulation.
Dans la région médiane et dorsale de l'embryon, l'action de Shh permet l'inhibition immédiate du facteur de transcription Pax7, l'activation du facteur Olig2 et, dans un second temps, l'activation du facteur de transcription Nkx2.2 qui va alors inhiber Olig2. Logiquement, la réponse à l'action de Shh est fonction du gradient de concentration de la protéine et est donc moins forte lorsque la concentration de Shh est plus faible. Néanmoins, il a été découvert que la réponse des facteurs de transcription peut également être influencée par la durée d'exposition à Shh. Ainsi, un réponse identique peut être observée face à une faible concentration mais à une période d'exposition à Shh longue ou au contraire face à une forte concentration mais une période d'exposition proportionnellement plus courte[2].
Remodelage et Evolution
Pour qu'une modification du patron de développement ait un rôle dans l'évolution, il faut nécessairement qu'elle soit héréditaire et qu'elle apporte un avantage sélectif à l'individu en améliorant sa fitness.
Tous les types de remodelage n'ont pas forcément la même probabilité de changer la direction de l'évolution. Les biais de développement et la sélection naturelle sont les principaux facteurs susceptibles d'influencer l'importance et la fréquence de ces différents mécanismes dans l'évolution. Par exemple, les mutations affectant les régions régulatrices des gènes sont plus fréquentes dans l'évolution que les mutations influençant les régions codantes. En effet les mutations sur les régions régulatrices n'affectent qu'une des fonctions d'un gène (changement de régulation) et non pas la totalité de ses fonctions (altération de la protéine synthétisée), elle a donc plus de chance d'être avantageuse.
Pour que la plasticité développementale influence la direction de l'évolution, il faut qu'il y ait une assimilation génétique. Cette dernière s'opère selon trois facteurs : il faut qu'il y ait un changement environnemental puis que celui-ci change le patron de développement (selon les biais du développement). Il faut ensuite que ce changement influence la fitness de l'individu, selon que l'influence est positive ou négative, les conséquences sur la direction de l'évolution seront différentes[2] - [3] - [1].
Références et Bibliographie
Références
- Alan C.Love Conceptual Change in Biology
- Evolution a Developmental approach Wallace Arthur
- Developmental Biology Scott F. Gilbert 2014
- Evolutionary Developmental Biology Brian K.Hall
- Rudolf A. Raff and Gregory, A.Wray Heterochrony: Developmental mechanisms and evolutionary results, 1989
- Scott. F. Gilbert Developmental Biology 2006
- Abzhanov, Arhat; Protas, Meredith; Grant, B. Rosemary; Grant, Peter R.; Tabin, Clifford J. (September 3, 2004), "Bmp4 and Morphological Variation of Beaks in Darwin's Finches"
- Abzhanov, Arhat; Kuo, Winston P.; Hartmann, Christine; Grant, B. Rosemary; Grant, Peter R.; Tabin, Clifford J. (August 3, 2006), "The calmodulin pathway and evolution of elongated beak morphology in Darwin's finches"
Bibliographie
- (en) Wallace Arthur, Evolution : A Developmental Approach, Wiley-Blackwell,
- (en) Brian K. Hall, Evolutionary Developmental Biology, Chapman and Hall,
- (en) Scot F. Gilbert, Developmental Biology : tenth edition,
- (en) Scot F. Gilbert, Developmental Biology : eighth edition,
- (en) Brian K.Hall, Keywords and Concepts in Evolutionary Developmental Biology,
- Mark Ridley, Evolution Biologique,
- (en) Cold Spring Harbor Symposium, Evolution : the Molecular Landscape
- (en) Alan C.Love, Conceptual Change in Biology, Springer,