Biais de développement
Le biais de développement est caractérisé par la production différentielle de phénotypes (caractères apparents d’un individu) à partir d’une variation génétique uniforme. Comme le processus de développement n’est pas linéaire, certains phénotypes sont plus probables, donc produits plus souvent, tandis que d’autres ne sont tout simplement pas produits. Il y a ainsi une distribution phénotypique non uniforme à partir des génotypes (information génétique d’un individu) qui eux, sont distribués uniformément. Le biais de développement est le résultat d’une hiérarchie et non d’un manque de variété des génotypes[1].
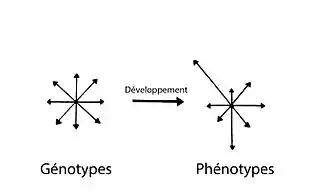
Il joue un rôle déterminant dans la direction que prend l’évolution. En effet, les différentes trajectoires phénotypiques empruntées durant l’évolution sont influencées par la facilité d’accès à certains phénotypes par rapport à d’autres, causée par le biais de développement. Étant donné que la sélection naturelle ne peut agir que sur les phénotypes produits par le processus de développement, la direction prise par les changements évolutifs est affectée par ce biais[1].
Donc, la sélection naturelle et le biais de développement interagissent. C’est cette interaction, et non un des deux processus par lui-même, qui détermine la direction que prend l’évolution[1].
Histoire
Le rôle que joue le biais de développement dans la direction de l’évolution a longtemps été controversé[2].
Charles Darwin reconnaissait qu’il n’y avait pas que la sélection naturelle pour déterminer les changements évolutifs, bien qu’elle en soit le moyen principal. Il croyait en la dépendance des caractères différents entre eux et sur la connectivité des différentes parties du corps lors du développement, comme c’est le cas pour les membres antérieurs et postérieurs. De son côté, Alfred Russel Wallace croyait plutôt à l’indépendance des caractères entre eux et non à une corrélation. Il avançait que les parties ou les organes du corps varient indépendamment des autres[2].
Bien que le concept du biais de développement ait commencé à être l’objet de controverses il y a longtemps, ce n’est que depuis les deux dernières décennies que des analyses se sont réellement penchées sur le sujet à travers la biologie évolutive du développement (évo-dévo). L’Évo-dévo a mené à des avancées majeures dans la compréhension des processus de développement et comment ils expliquent les modèles de la diversification évolutive[3].
Effets des mutations
Les caractères morphologiques ne sont pas déterminés par l’absence ou la présence d’un gène, mais plutôt par les changements qui occurrent dans les mécanismes de régulation, qui affectent l’endroit et le moment où le gène est produit[4].
Lorsqu’il y a des mutations, dans la séquence ADN, il peut y avoir des changements phénotypiques. Le biais de développement est particulièrement sensible aux mutations. Même de petits changements du système régulateur causés par des mutations neutres, se situant dans les introns (la partie non codante du gène), ont un effet sur le modèle du biais de développement. À long terme les mutations affectant le génome peuvent finir par modifier les phénotypes accessibles. Les phénotypes qui seront présents chez les générations futures dépendent fortement des génotypes des générations actuelles. Les mutations dans ces génotypes vont dicter les prochains phénotypes accessibles[1].
Le biais de développement évolue aussi, ce qui génère de la variété dans l’évolutivité (capacité de varier en phénotypes accessibles au cours du temps)[1].
Effets du biais de développement
Le fait qu’il y ait un biais dans la production des différents phénotypes peut aussi orienter les réponses des organismes qui font face à de nouveaux environnements. Le biais de développement peut entraîner une spéciation (formation de nouvelles espèces) plus rapidement à l’aide des lignes de moindre résistance, où les phénotypes sont plus faciles d’accès et où les directions alternatives de l’évolution sont moins probables. Il y a plusieurs exemples d’évolution parallèle, comme chez les poissons de la famille des Cichlidés en Afrique, où des espèces indépendantes développent des caractéristiques similaires. Ces exemples suggèrent que la façon dont les variations phénotypiques sont générées oriente l’évolution adaptative selon certaines trajectoires [4]
Le biais de développement peut aussi avoir un effet tampon, dans le sens qu’il peut empêcher la production de phénotypes nuisibles pour la survie d’un organisme. En effet, un biais dans le processus de développement du système neural, qui est responsable des comportements de l’animal, peut faire en sorte que les phénotypes néfastes, causant des comportements mal exécutés ou l’absence d’un comportement nécessaire à la survie, ne soient pas produits[1].
Dans le même sens, le biais de développement peut aussi ouvrir des opportunités pour de nouveaux changements évolutifs en conduisant à des phénotypes responsables de comportements avantageux et donnant une plus grande fitness (succès reproducteur d’un individu)[1].
Biais global et local
Le biais global est un biais propre au système de développement. Il représente les phénotypes produits par un échantillon de plusieurs génotypes aléatoires. Il montre une dégénérescence du système développemental, c’est-à-dire la production du même phénotype par plusieurs génotypes différents. Étant donné que les génotypes mènent au même phénotype, cette dégénérescence est phénotypiquement neutre. Or, cette dégénérescence n’a pas les mêmes modèles de biais locaux[1].
Le biais local est un biais de développement génotypiquement dépendant. Il varie énormément selon les changements des génotypes, même ceux qui sont phénotypiquement neutres. Il dépend de la dynamique du processus développemental et non de l’ensemble des mécanismes présents dans le système de développement. Cela fait en sorte que les modèles du biais peuvent changer drastiquement lors de l’évolution, par l’accumulation de mutations, neutres ou non. C’est le biais local qui est responsable de la production différentielle de phénotypes à partir d’une distribution uniforme de génotypes. Il est déterminé en développant chaque phénotype mutant, obtenu en performant toutes les substitutions de base individuelle sur un génotype, et en calculant la fraction des mutations donnant le même phénotype[1].
Au cours de l’évolution, la dépendance locale du biais de développement dicte les transformations phénotypiques qui auront lieu au sein des populations. Le modèle du biais local pour un certain génotype est différent des autres biais locaux et du biais global. Ces différences entre les biais locaux montrent qu’ils dépendent plus des interactions de développement d’un génotype en particulier. Étant donné que les phénotypes accessibles sont aussi dépendants localement, les génotypes de la population actuelle vont déterminer les phénotypes présents dans la génération future. Un certain trajet évolutif emprunté par une population dépend fortement des biais locaux de chaque individu. La neutralité phénotypique de la dégénérescence peut faire en sorte que les mutations ne changeant pas le phénotype s’accumulent graduellement et changent peu à peu le contexte développemental. Ce qui pourrait permettre à de nouveaux phénotypes d’être accessibles. Certains génotypes pourraient finir par donner un phénotype ayant un avantage pour les individus, grâce au biais local[1].
Exemple des segments des centipèdes
Les centipèdes font partie d’une des classes des myriapodes (mille-pattes). Le nombre de segments portant des pattes chez les centipèdes est un exemple clair du biais de développement. La distribution du nombre de ces segments dans les centipèdes montre des caractères impairs allant de 15 à 191 segments selon les espèces. Or, il n’y a aucune des quelque 3000 espèces de centipèdes qui possède un nombre pair de segments[5].
.jpg.webp)

L’ordre Lithobiomorpha regroupe environ 1100 espèces ayant 15 segments porteurs de pattes à l’âge adulte. Par contre, lorsqu’ils naissent, ils possèdent moins de segments et continuent d’en ajouter durant leur développement jusqu’à l’âge adulte. Il y a donc des stades où les lithobiomorphes ont un nombre pair de segments de tronc, mais seulement à l’état juvénile. Pour ce qui est de l’ordre Geophilomorpha, regroupant plus de 1000 espèces, ils possèdent un nombre beaucoup plus élevé et varié de segments qui portent des pattes que les lithobiomorphes. Or, contrairement à ces derniers, les géophilomorphes possèdent tous leurs segments à leur naissance[2].
Le gène de polarité segmentaire engrailed est impliqué dans la segmentation des centipèdes. Le patron d’expression de ce gène forme des bandes à la partie postérieure de chaque segment. Il a été montré que chaque bande apparaît une à la fois et non par paire. On serait tenté de croire que ce processus pourrait s’arrêter après n’importe laquelle de ces bandes, formant ainsi un nombre aléatoire de segments. Le dernier segment de tronc pourrait aussi être supprimé par apoptose, comme pour les tissus de la région interdigitale des tétrapodes. Or, ce n’est pas le cas[2].
Ce phénomène est un exemple de biais de développement, plus précisément de biais global, car un processus de développement pousse vers la production de segments impairs, alors que la production de segments pairs subit une contrainte absolue. Cela ne veut pas dire que le caractère n’est pas présent car il n’est pas adaptatif, mais bien parce qu’il ne peut pas exister en raison de mécanismes développementaux[6]. Une forme de sélection encore inconnue qui agirait contre la production de segments impair est très improbable[2].
Articles connexes
Notes et références
- (en) Psujek, S. Beer, R. D. Developmental bias in evolution: evolutionary accessibility of phenotypes in a model evo-devo system, 2008, Evolution & Development (Vol. 10, No 3), p. 375-390
- (en) Wallace, A. The interaction between developmental bias and natural selection: from centipede segments to a general hypothesis, 2002, Heredity, p. 239-246
- (en) Brakefield, M. P. Evo-devo and accounting for Darwin's endless forms, 2011, Phylosophical transactions of the royal society (Vol 366), p. 2069-2075
- (en) Brakefield, M. P. Evo-devo and constraints on selection, 2006, TRENDS in ecology and evolution (Vol. 21, No 7), p. 362-368
- (en) Chipman, D.A. Wallace, A. Akam, M. A Double Segment Periodicity Underlies Segment Generation in Centipede Development, 2004, Current Biology (Vol 14), p. 1250-1255
- (en) Damen, M. G. W. Arthropod Segmentation: Why Centipedes Are Odd, 2004, Current Biology (Vol 14), R557-R559