Muscle lisse
Le muscle lisse est un type de muscle dont, à l'inverse du muscle strié, les protéines contractiles ne sont pas organisées sous forme de sarcomères, ce qui lui ôte l'aspect strié lors de l'observation sous microscope. Le muscle lisse se contracte lentement et avec moins de force que le muscle strié. Il est sous le contrôle du système nerveux autonome et du système endocrinien et fonctionne donc de manière indépendante de la volonté. Il joue ainsi un rôle majeur dans la vie végétative, c'est-à-dire qu'il est essentiel à la mise en place et à l'exécution de fonctions vitales à un organisme. Le tissu musculaire lisse est pour cette raison retrouvé dans la couche musculeuse de la paroi d'organes creux des systèmes digestif, respiratoire, circulatoire, urinaire et génital, et procurent à la paroi de ces organes ses capacités contractiles. Le muscle lisse est composé de cellules musculaires nommées cellules musculaires lisses ou plus rarement léiomyocytes baignant dans un gel de matrice extracellulaire.
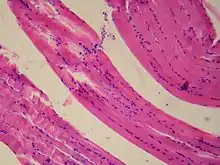
Description des cellules musculaires lisses
Les cellules musculaires lisses sont des cellules fusiformes uninucléées mesurant entre 20 et 200 µm. Elles ont des extrémités pointues et bifides. Leur noyau est bien visible, en général central et allongé, et situé dans la partie la plus large de la cellule. Le cytoplasme apparaît assez homogène. Le réticulum sarcoplasmique n'a pas la régularité de celui des fibres musculaires striées. On distingue à l'intérieur des myofibrilles (sous forme de myofilaments) orientées selon le grand axe de la cellule et composées comme pour tout muscle de 2 types de filaments constitués de protéines contractiles : les filaments fins d'actine et les épais de myosine. Les myofilaments fins entourent un myofilament épais, et la contraction a lieu en présence d'ATP. La calmoduline joue également un rôle très important dans la contraction de ces cellules. Les cellules musculaires lisses sont également constituées de filaments intermédiaires de desmine et de vimentine.
On retrouve comme structures particulières :
- des corps denses, équivalent des stries Z (alpha-actinine) des sarcomères ;
- des structures membranaires particulière nommées « cavéoles », en lien avec la cavéoline (protéine lié au phénomène d'endocytose), qui jouent un rôle proche des tubules T et régulent le taux de Ca2+ intracellulaire ;
- des plaques d’adhérence qui font le lien entre les filaments d’actine et la matrice extracellulaire (la fibronectine de la MEC) via des intégrines.
Du point de vue biochimique, les filaments d'actine sont constitués d'α-actine du muscle lisse, qui est une isoforme que l'on retrouve relativement spécifiquement dans le muscle lisse, tandis que les filaments de myosine sont constitués de chaîne lourde de la myosine du muscle lisse qui est une isoforme strictement spécifique du muscle lisse. Ces deux protéines sont utilisées comme marqueurs histologiques du muscle lisse. D'autres protéines du cytosquelette des cellules musculaires lisses sont également utilisés comme marqueurs histologiques telles que la desmine (que l'on retrouve aussi dans les muscle cardiaque et squelettique) et la forme lourde de la caldesmone.
Les cellules baignent dans une substance hyaline comportant des cloisons pellucides. Elles sont organisées en faisceaux formant différentes couches et entourés de tissu conjonctif lâche (fibres dites anastomosées irrégulières). Ces fibres constituent des unités fonctionnelles contractiles disposées parallèlement les unes aux autres. Les cellules sont placées de telle sorte que la partie renflée (médiane) d'une cellule vienne en contact avec les extrémités effilées des cellules adjacentes. La zone de jonction constitue le nexus (où les membranes plasmiques des cellules s'unissent).
En culture, in vitro, les cellules musculaires lisses ont la particularité de se dédifférencier rapidement pour tendre vers un phénotype proche des cellules mésenchymateuses. Elles perdent alors leur capacité d'inhibition de contact et se mettent à proliférer et à s'accumuler en plusieurs couches par endroits ce qui leur confère la caractéristique visuelle au microscope de culture en « collines et vallées ». Ce phénomène de dédifférenciation s'accompagne de la modification d'expression de marqueurs histologiques, telles que la perte d'expression de l'α-actine du muscle lisse et de la chaîne lourde de la myosine du muscle lisse au profit d'isoformes comme la chaîne lourde de la myosine non-musculaire pour les filaments de myosine[1]. Ce processus de dédifférenciation est retrouvé in vivo dans des processus pathologiques telles que l'athérosclérose et la resténose post-angioplastie.
On retrouve ces cellules sous différentes formes chez les animaux, notamment chez les humains. On peut les observer :
- à l'état isolé dans des capsules, le scrotum, le mamelon… ;
- sous forme de muscle lisse pouvant être :
- en couches dans les vaisseaux, le tube digestif…,
- en muscles organisés comme pour les muscles arrecteur du poil, dans l'iris.
Description du muscle lisse
Les muscles lisses multiunitaires sont innervés comme les muscles striés, avec une synapse par cellule.
Les muscles lisses unitaires (comme les viscères) ont une innervation pour un groupe de cellules particulières. Ces cellules particulières sont douées d'automaticité, transmettant le potentiel de membrane instable aux autres cellules lisses. L'innervation du système nerveux végétatif sert à moduler la contraction musculaire automatique.
Les muscles lisses sont sous le contrôle du système nerveux autonome parasympathique dont le neurotransmetteur est l'acétylcholine et orthosympathique viscéral dont les neurotransmetteurs sont l'acétylcholine (au niveau de la jonction neuro-neuronale) et la noradrénaline (à la jonction avec le muscle lisse).
Localisation du muscle lisse
Les muscles lisses se retrouvent dans les parois, nommées tuniques, des :
- parois vasculaires : dans la paroi des artères où ils sont impliqués dans la tonicité artérielle et la vasomotricité ;
- voies digestives : dans les parois du tube digestif, au niveau de la musculeuse, constituée d'une couche musculaire externe longitudinale (dont la contraction raccourcit un segment du tube digestif) et d'une couche musculaire circulaire interne (qui diminue le diamètre de la lumière ; les sphincters ne sont que des épaississements de cette circulaire interne) où ils assurent la progression du bol intestinal par péristaltisme ;
- voies respiratoires : par exemple dans les bronches ;
- voies urinaires : par exemple dans la vessie ;
- voies génitales : par exemple dans l'utérus ;
- globes oculaires (muscle ciliaire) et de l'iris.
Fonction
Le muscle lisse a pour fonction d'aider au transport de différents milieux à l'intérieur de l'organisme. Ces différents milieux sont :
- le sang pour le muscle lisse des vaisseaux sanguins ;
- de l'air pour le muscle lisse des bronches ;
- de la nourriture pour le muscle lisse du tube digestif ;
- de l'urine pour le muscle lisse des reins, de la vessie et des faisceaux transportant l'urine ;
- du derme de la peau ;
- le produit de sécrétion pour les cellules myoépithéliales (cellules musculaires lisses) des glandes exocrines.
Le monoxyde d'azote endogène est un médiateur qui a pour effet de les relâcher. Une forte concentration de ce gaz peut produire une intoxication.
Différences avec les muscles squelettiques
Les muscles lisses fonctionnent inconsciemment, ils sont contrôlés par les systèmes ortho- et parasympathiques, tandis que les muscles squelettiques provoquent des mouvements volontaires sous le contrôle des nerfs moteurs.
Les muscles squelettiques apparaissent striés à l'observation sous microscope alors que les muscles lisses ne l'apparaissent pas.
Les muscles lisses possèdent moins de myosine que les muscles squelettiques.
La forme en fuseau des cellules musculaires lisses est beaucoup plus petite que celle des muscles squelettiques. En effet, les fibres musculaires squelettiques ont un diamètre qui varie entre 10 et 100 μm et une longueur qui peut atteindre plusieurs dizaines de centimètres alors que les cellules musculaires lisses ont une largeur comprise entre 2 et 10 μm et une longueur variant de 50 à 400 μm.
La force de contraction d’un muscle lisse est moins importante que celle d’un muscle squelettique, mais un muscle lisse peut rester contracté pendant une période plus longue.
Les muscles cardiaques ressemblent aux muscles squelettiques bien qu’ils fonctionnent inconsciemment. En effet, ils présentent des stries lors de l'observation microscopique comme le muscle squelettique mais sont contrôlés par le système nerveux orthosympathique comme le muscle lisse.
Physiologie[2]
Mécanisme de contraction
Les concentrations intracellulaires de Ca2+ augmentent lorsque :
- du calcium d'origine extracellulaire entre dans la cellule, par des canaux calciques chimio-dépendants, et voltage-dépendants ;
- du calcium est libéré par le réticulum endoplasmique.
Ce Ca2+ va se lier à la calmoduline (CaM), de telle sorte que quatre ions soient fixés sur chaque molécule de calmoduline.
Le complexe Ca2+-calmoduline va ensuite activer la kinase de la chaîne légère de myosine (MLCK).
La MLCK va ainsi pouvoir phosphoryler les chaînes légères des têtes de myosines et ipso facto, augmenter l'activité de la myosine ATPase.
La myosine ainsi activée, les ponts de myosine actifs vont pouvoir glisser le long de l'actine et développer une tension musculaire.
Mécanisme de relaxation
Le Ca2+ libre dans le cytosol diminue quand :
- le Ca2+ est transporté hors de la cellule, en passant à travers la membrane par un antiport Na+/Ca2+ : la sortie d'un ion Ca2+ contre son gradient électrochimique, s'effectue en même temps que l'entrée de deux ions Na+ dans le sens de leur gradient électrochimique. Le Ca2+ peut aussi être pompé hors de la cellule grâce aux pompes calciques (ATPases) ;
- le Ca2+ retourne dans le réticulum endoplasmique grâce aux pompes calciques à activité ATPases (SERCA), qui assurent un transport actif de Ca2+ en utilisant de l'ATP. Une fois dans le réticulum, le calcium y est maintenu par la séquestrine.
Le Ca2+ se détache ensuite de la calmoduline (CaM), puis la myosine phosphatase détache le phosphate de la myosine, ce qui diminue l'activité de la myosine ATPase. Cette diminution progressive d'activité conduit à la diminution de la tension musculaire, à la relaxation musculaire.
Importance biologique des contractions rythmiques
Remarquons tout d’abord que les contractions musculaires du gésier sont spontanées. Elles sont donc visibles en l’absence de stimulations.
Le gésier est soumis à deux contractions différentes qui se font l’une à la suite de l’autre. L’une est longitudinale et l’autre est circulaire. Les couches de muscles lisses longitudinales permettent le raccourcissement et la dilatation de l’organe tandis que les couches circulaires permettent la variation du diamètre de la lumière de l’organe. Cette succession entre ces deux types de contractions permet le péristaltisme et donc l’évacuation la quantité de nourriture ingérée par le ver de terre.
Vitesse de régularisation
Avant de commencer l'expérience, le ver est recouvert d'éthanol afin d'être anesthésié. Au début, la membrane est recouverte d'éthanol, ce qui inhibe le système. Il faut attendre que l'éthanol se soit dissipé pour observer les contractions « normales » du gésier, c'est-à-dire les contractions qu'aurait le gésier s'il était toujours à l'intérieur du ver de terre.
Si le gésier était placé exactement à la verticale, nous apercevrions uniquement les contractions longitudinales du gésier. Dans notre expérience, le gésier n'est pas parfaitement perpendiculaire au sol ; nous apercevons dès lors de petites contractions circulaires (petits pics) qui suivent les contractions longitudinales (grands pics). Vu que le gésier est un peu en oblique, les pics mesurés correspondant aux contractions longitudinales sont d'amplitudes plus faibles qu'ils ne le sont en réalité.
Effet de la température
On observe trois cas de température. Elle est initialement à une température ambiante de 22 °C, puis à une température de 11 °C, ensuite à une température de 33 °C et retourne après à une température de 22 °C. Il est alors possible d'observer que l'amplitude de la force des contractions est relativement équivalente quelle que soit la température. Par contre, la fréquence varie en fonction de la température.
Lorsque la température augmente, la fréquence des contractions augmente également. Les contractions sont provoquées par le passage d'ions à travers la membrane. Or une augmentation de la température entraîne une agitation des ions et donc une augmentation de l'énergie cinétique de ces derniers. Les ions passent donc plus rapidement à travers la membrane. Par conséquent, la fréquence des contractions est augmentée, ce qui se traduit par le rapprochement des pics.
À l'inverse, une diminution de la température entraîne une diminution de l'activité cinétique des ions. Les ions prennent plus de temps pour traverser la membrane, par conséquent, la fréquence des contractions est plus faible.
De plus, lorsque la température est froide, l'amplitude maximale augmente légèrement et atteint 0,07 N. Cela est également dû à la cinétique des ions. En effet, le calcium va prendre plus de temps pour sortir et donc il restera du calcium lorsque la prochaine contraction va se produire. Celle-ci sera plus forte, car il y aura une plus grande quantité de calcium à l'intérieur de la cellule. La force contractile ne redescend jamais en dessous de 0,045 N, ce qui montre bien qu'une quantité de calcium réside continuellement à l'intérieur de la cellule. Lorsque la température est trop basse, le muscle n'est donc jamais relâché complètement ; il reste continuellement contracté.
Nous pouvons remarquer qu'à une température chaude de 33 °C, la fréquence est élevée au début puis elle diminue progressivement au fur et à mesure de l'expérience. Cela illustre le phénomène de fatigue musculaire. Pour effectuer une contraction, de nombreux ions doivent traverser la membrane. Ce déplacement d'ions demande beaucoup d'énergie sous forme d'ATP. À un certain moment, il n'y a plus assez d'ATP pour assurer une fréquence de contraction élevée même à haute température et cela se traduit par une diminution de la fréquence des contractions.
Effet de la concentration en K+
Lorsque la concentration extérieure de K+ est intensifiée, la plus grande différence que l'on peut observée est l'augmentation de la fréquence de battements, qui varie du quitte au double. Par comparaison, la variation d'amplitude des pics est négligeable, étant selon les valeurs mesurée de 15 % en moyenne, mais diminuant clairement avec le temps, au contraire de la fréquence.
Le K+ est responsable d'une grande partie de la différence de potentiel entre l'intérieur et l'extérieur de la cellule, étant beaucoup plus concentré dans la cellule. Par conséquent, changer sa concentration dans le milieu extérieur revient à changer le potentiel de repos. Selon l'équation \ref{eq:potential}, on voit facilement qu'augmenter la concentration extérieure en potassium, comme le logarithme est une fonction croissante, fait augmenter la différence de potentiel. En d'autres termes, puisque cette différence est négative, on diminue la valeur absolue de la différence de potentiel de repos et cela la rapproche du seuil de déclenchement d'un potentiel d'action. Un potentiel de repos plus proche du seuil se traduit par un retour plus rapide à l'équilibre et une augmentation de la fréquence de battement du muscle.
![{\displaystyle V={\frac {RT}{F}}\ln {\frac {P_{K}\left[K_{out}^{+}\right]+P_{Na}\left[Na_{out}^{+}\right]+P_{Cl}\left[Cl_{in}^{-}\right]}{P_{K}\left[K_{in}^{+}\right]+P_{Na}\left[Na_{in}^{+}\right]+P_{Cl}\left[Cl_{out}^{-}\right]}}}](https://img.franco.wiki/i/9d0a8fcfe0209ab613fe75aa882eb9eded6ae265.svg)
Ce changement du potentiel de repos est identique pour tous les muscles, un muscle squelettique ou cardiaque verrait donc aussi sa fréquence augmentée.
Effet de la concentration en Na+
Selon l'équation juste au-dessus, on pourrait penser qu'augmenter la concentration en sodium extérieur aurait le même effet que pour le potassium, cependant le terme de perméabilité au sodium est bien plus faible que celui du potassium. Par conséquent, l'effet d'un changement de concentration en Na+ serait 50 fois moins important (PNa+=0.02 si PK+=1), et en deviendrait négligeable.
Notes et références
- (en) Gary K Owens, « Molecular control of vascular smooth muscle cell differentiation », Physiological Reviews, vol. 75, , p. 487-517 (lire en ligne)
- Cours de Physiologie d'Anh Tuan Dinh-Xuan