Microcystine
Le terme microcystine désigne une famille de toxines produites par différents genres de cyanobactéries. On désigne les microcystines comme des cyanotoxines dans la mesure où elles représentent une famille de composés produits par les cyanobactéries qui présente un effet délétère sur l'activité biologique d'un organisme dans un environnement donné[1]. Les cyanobactéries productrices de microcystine ont principalement été identifiées en eau douce. Cependant, en milieux marins certains genres de cyanobactéries tels que Geitlerinema ou Synechococcus ont également été décrit comme producteurs de microcystines[2]. En 2004, la présence de microcystines en milieux terrestres a également pu être mise en évidence. Les microcystines seraient également produites par des cyanobactéries du genre Nostoc évoluant en symbiose avec certains lichens[3].
Les microcystines sont à ce jour les cyanotoxines les mieux caractérisées et les plus répandues au sein du phylum des cyanobactéries[4]. Cette molécule, très résistante, est dite « heptapeptide » car constituée de sept acides aminés qui se rejoignent aux extrémités pour former une structure cyclique.
Les pullulations (sous forme d'efflorescences planctoniques le plus souvent) de cyanobactéries produisant de la microcystine semblent en augmentation depuis quelques décennies, en raison de l'eutrophisation générale de l'environnement et elles pourraient être sous-estimées.
Elles causent la mort d'animaux sauvages et domestiques (bétail, chiens, chats, etc.)[5] et peuvent aussi affecter les humains[6] (ex. : soixante personnes mortes en 1996 lors d'une intoxication à l'hôpital de Caruaru au Brésil[7]).
Les microcystines sont le type de cyanotoxine le plus étudié à ce jour. En France elles sont notamment suivies en Bretagne depuis 1995 où leur présence a été détectée dans près de 65 % des sites étudiés[8] - [9]
En 2004, les toxicologues et biologistes en avaient déjà identifié plus de soixante-dix variantes[10]. Elles sont produites par plusieurs genres de cyanobactéries qui regroupent de nombreuses espèces présentes sur tous les continents où elles produisent occasionnellement (et de plus en plus souvent – chaque année dans certains lacs) des efflorescences potentiellement hépatotoxiques. Ces bactéries appartiennent principalement aux genres Oscillatoria, Nodularia, Microcystis, Planktothrix, Anabaena et Aphanizomenon. Une même efflorescence peut associer plusieurs espèces appartenant à plusieurs de ces genres de bactéries et leurs toxines peuvent avoir des effets synergiques.
Enjeux
Ils sont toxicologiques et sanitaires (qualité des eaux de baignade, qualité de l'eau potable[11] - [12] - [13], qualité de la viande de gibier ou du bétail, et même du lait dans lequel la toxine peut passer (chez la vache par exemple[14])), mais aussi écotoxicologiques et de connaissance, et relèvent notamment de l'évaluation environnementale, de la surveillance environnementale[15] et de l'évaluation et la gestion des risques[16].
Biochimie, description
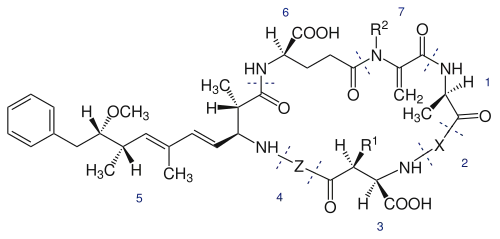


Squelette de la microcystine : 1, D-Alanine ; 2, X (variable) ; 3, D-MeAsp (D-érythro-b-méthyl-D-acide aspartique) ; 4, Z (variable) ; 5, Adda (acide (2S, 3S, 8S, 9S)-3-amino-9-méthoxy-2-6-8-triméthyl-10-phényldéca-4,6-diènoïque) ; 6, D-glutamate ; 7, Mdha (N-méthyldéshydroalanine). R1 et R2 sont H (déméthylmicrocystines) ou CH3[17].
Variantes de la toxine
Elles proviennent majoritairement de substitution d’acide aminé en position 2 et 4 ou bien du retrait/ajout d’un groupe méthyle sur les fonctionnalités 3 et 7.
Les acides aminés les plus communs à la position X sont la leucine (L), l’arginine (R) et la tyrosine (Y), alors qu’à la position Z on retrouve généralement l’arginine (R), l’alanine (A) et la méthionine (M)[10]. Le tableau suivant présente les différents acides aminés retrouvés à chaque position. La combinaison de ces possibilités explique le fait qu’il y ait autant de microcystines.
| AA 1 | AA 2 | AA 3 | AA 4 | AA 5 | AA 6 | AA 7 |
|---|---|---|---|---|---|---|
| D-Ala | L-Leu | D-MeAsp | L-Arg | Adda | D-Glu | Mdha |
| D-Ser | L-Arg | D-Asp | L-Aib | ADMAdda | D-MeGlu | Dha |
| L-Ala | L-Ala | DMAdda | E(OMe) | Dhb | ||
| L-Glu | L-Glu | (6Z)Adda | L-Ala | |||
| L-GluMe | L-GluMe | L-MeSer | ||||
| L-Har | L-Har | L-Ser | ||||
| L-Hil | L- Hph | Mdhb | ||||
| L-Hph | L-Hty | MeLan | ||||
| L-Hty | L-Leu | |||||
| L-Met | L-Met | |||||
| L-Met(O) | L-Met(O) | |||||
| L-Phe | L-Phe | |||||
| L-ThTyr | L-Trp | |||||
| L-Trp | L-Tyr | |||||
| L-Tyr | L-Val |
Aib = Acide aminoisobutytique
ADMAdda = O-Acétyl-O-déméthylAdda
Dha = Déshydroalanine
Dhb = Déshydrobutyrine
DMAdda = O-DéméthylAdda
E(OMe) = Acide glutamique méthylester
Har = Homoarginine
Hil = Homoisoleucine
Hph = Homophénylalanine
Hty = Homotyrosine
MeLan = N-Méthylanthionine
Met(O) = Méthionine-S-oxyde
MeSer = N-Méthylsérine
ThTyr = 1,2,3,4-Tétrahydrotyrosine
(6Z)Adda = Stéréoisomère de Adda
Selon leur composition, les microcystines présentent une gamme de poids moléculaires située entre 900 et 1 100 Da.
Ces molécules sont très solubles dans l’eau et peu volatiles.
Diversité des organismes producteurs
Il existe une importante diversité d'organismes producteurs de microcystines.
Résistance
À cause de leurs structures cycliques, elles sont très résistantes face aux conditions ambiantes et elles tolèrent différentes conditions de pH et de températures. De plus, elles peuvent rester stables pendant des années[19], voire sous forme de traces durant des siècles, millénaires[7] voire centaines de milliers d'années si elles ne sont pas en contact avec la lumière.
Prévalence
Bien que la première mort par empoisonnement bactérien de groupes d'animaux n'ait été rapportée qu'au XIXe siècle (par Georges Francis en 1878 dans le journal Nature[20]), des épidémies zoonotiques de ce type ont été rapportées sur tous les continents au XXe siècle, préoccupant en raison de l'augmentation estimée des blooms saisonniers dans de nombreux pays, et de nombreux indices laissent penser que de tels phénomènes sont plus anciens encore ; les populations indigènes amérindiennes du Nord de l'Amérique, d'Afrique et d'Australie connaissaient la nature toxique de certaines pullulations de cyanobactéries[21].
Ainsi, des analyses paléobiochimiques de sédiments anciens composés de cyanobactéries très probablement décomposés par d'autres bactéries, benthiques montrent ce qui semble être des pigments et des toxines caractéristiques des cyanobactéries. Ceci laisse penser que des pullulations de cyanobactéries toxiques ont autrefois périodiquement eu lieu de manière saisonnière, par exemple dans des lacs de cratères volcaniques où l'on trouve des accumulations inhabituelles de cadavres de vertébrés aujourd'hui fossilisés (par exemple à Neumark-Nord ou dans d'autres sédiments lacustres tertiaires tels que le schiste bitumineux de Messel (ancien lac volcanique)), y compris des mammifères[22]. La position de certains squelettes laisse penser que leurs propriétaires aient pu mourir empoisonnés[23].
En France
Dans ce pays, les premiers signalements de blooms planctoniques à cyanophycées dans les rivières, mares et réservoirs d'eau potable (lacs de barrages) ont été effectués en Bretagne au milieu des années 1990, avec 25 réservoirs identifiés, le plus souvent colonisés par la cyanobactérie Microcystis aeruginosa[24].
La cyanophycée filamenteuse Planktothrix agardhii a par exemple été trouvée dans un lac peu profond et eutrophe, utilisé pour les loisirs et la pêche et relié à la Seine par un canal artificiel, avec dans ce cas une biomasse élevée toute l'année, même en hiver (7,9 × 104 à 4,5 × 106 trichomes/L)[25]. était dans ce cas associé à une autre cyanobactérie (Limnothrix redekei) toutefois moins présente. Les analyses ont révélé la présence de cinq microcystines différentes dans ces eaux.
Toxicité
Toxicité pour l'humain
Les variants les plus toxiques le sont autant que le curare (à dose égale)[7].
Des empoisonnements dus à la consommation d'eau contaminée par la cyanobactérie toxique Cylindrospermopsis raciborskii ont été étudiés à Palm Island (nord du Queensland, Australie)[26]. Sa toxine a été finement étudiée et même biosynthétisée in vitro, et sa détection et quantification par analyse chimique et biologique ont été améliorées[26]. Les facteurs favorisant la production de toxines par les isolats de culture de C. raciborskii sont mieux compris de même que ceux qui provoquent leur libération dans l'environnement. Diverses stratégies de gestion sont testées dans le monde visant à minimiser la production de toxine ou diminuer ses effets néfastes, par traitement de l'eau ou du milieu ou en cherchant à neutraliser la toxine ou à empêcher la prolifération des cyanophycées. On cherche encore à mieux comprendre les facteurs de risque, la génétique de ces espèces[26], mais des techniques moléculaires permettent maintenant de caractériser les souches « toxiques » et « non toxiques »[26].
Mécanisme de la toxicité
Les microcystines sont des molécules hépatotoxines (qui détruisent les cellules du foie en désorganisant leur cytosquelette).
Ce sont des inhibiteurs de protéines de types phosphatases (PP1 ou PP2A) qui exécutent la déphosphorylation de phosphoprotéines intracellulaires chez les eucaryotes. Cette inhibition entraîne un dérèglement de la structure de la cellule qui conduit à sa nécrose ; une nécrose massive d'hépatocytes entraîne une hémorragie interne et éventuellement la mort.
De plus, la phosphorylation excessive de protéines anti-tumorales fait en sorte que les microcystines sont considérées comme des promoteurs de tumeurs[17].
Toxicité variable et saisonnière
La toxicité de cette molécule varie fortement d'une part selon l'espèce qui est en contact avec la toxine (certaines espèces y sont naturellement plus résistantes), et d'autre part selon la forme (variant moléculaire) de la molécule de microcystine.
La toxicité des différentes variantes de la microcystine semble dépendre du degré de méthylation de certains acides aminés.
Lorsque l’entité ADDA est méthylée, la toxicité de la microcystine diminue considérablement, suggérant que cette partie de la molécule est le site d’interaction majeur avec les phosphatases[27]. De plus, la forme linéaire des microcystines est reconnue pour être cent fois moins toxique[17].
Une saisonnalité des mortalités par intoxication est observée et peut l'être rétroactivement par l'étude des sédiments.
Toxicité pour l'animal, et écoépidémiologie
Certains événements attirent l'attention par leur importance géographique (ex. : bloom observé sur 1 000 km environ sur la Darling en Australie, qui a tué au moins 2 000 bovins[7]) ou par une espèce qui semble particulièrement vulnérable (ex. : 320 alligators morts empoisonnés de 1997 à 2001 à la suite des blooms observés sur le lac Griffin (Floride)[7]). En France, en 2003, plusieurs chiens sont morts après avoir bu de l'eau de la Loue[7], dont les sources coulent sur une décharge de munitions non explosées datant de la première Guerre mondiale.
Les morts animales surviennent souvent durant mêmes saisons avec certaines conditions météorologiques identiques (pas de vent durant une certaine période[28]), aux mêmes endroits (mêmes points d'eau stagnante) et généralement quand les animaux n'ont pas d'accès à un point d'eau sain à proximité[28]. Mais les eaux contaminées par les toxines de certaines cyanophycées semblent attirer certains animaux, et en laboratoire, la souris se montre plus attirée par une eau contaminée que par une eau propre ou une eau contenant d'autres bactéries, buvant l'eau toxique jusqu'à en mourir[29]. Il est possible qu'une telle attraction « fatale »[28] se produise parfois dans la nature ou que des espèces n'aient pas su choisir entre deux points d'eau, l'un contaminé et l'autre ne le semblant pas (ex. : 15 chiens et 20 moutons morts au Royaume-Uni après avoir bu dans le Rutland water (Leicestershire en 1989[28] - [30])).
Certains oiseaux qui se nourrissent en filtrant l'eau (canards, flamants roses[31]…) y sont plus exposés.
Risques sanitaires
Ils sont liés au risque d'exposition en cas de :
- baignade ;
- boisson contaminée ou contamination de grands lacs-réservoirs destinés à produire de l'eau potable[32] ;
- inhalation d'aérosols ou embruns contenant la toxine[33] - [34] ;
- utilisation d'eau de surface contaminée pour l'irrigation ou l'arrosage de cultures ou jardins potagers[35] ;
- alimentation contaminée.
Les coquillages et organismes filtreurs peuvent bioconcentrer cette toxine (moule zébrée, anodontes…). De plus la moule zébrée résiste à de hautes doses de microcystine[36] et semble favoriser les cyanobactéries qui en produisent[37].
La présence de microcystines dans certaines microalgues utilisées comme complément alimentaire, en particulier l'algue Klamath, a conduit les autorités sanitaires de l'Oregon à imposer un seuil de microcystines à ne pas dépasser dans l'algue Klamath produite à destination humaine en 1996[38]. Cette algue continue à poser des problèmes selon des études italienne[39] et allemande[40] qui ont appelé en à remettre en question son autorisation de mise sur le marché.
Les blooms algaux sont de plus en plus fréquent en eaux intérieures (lacs de barrages, lacs et étangs urbains protégés du vent) et des contaminations aux microcystines sont problématiques, en Europe y compris[41].
Médecine légale ou vétérinaire
Une microcystine peut être évoquée quand un animal présente des symptômes d'empoisonnement ou qu'il meurt dans un contexte où un bloom bactérien de cyanophycées a été constaté. Des analyses biochimiques peuvent permettre de trouver des traces de ces toxines ou de la bactérie qui les produit dans l'estomac. Une nécropsie pratiquée lors d'une autopsie précoce montre une attaque du foie avec nécroses cellulaires. Une étude plus poussée permet de trouver des traces de la toxine dans les tissus.
Notes et références
- (en) Ecology of Cyanobacteria II : Their Diversity in Space and Time, Dordrecht, Springer Netherlands, (ISBN 978-94-007-3854-6 et 9789400738553, DOI 10.1007/978-94-007-3855-3, lire en ligne)
- (en) Franca M. Buratti, Maura Manganelli, Susanna Vichi et Mara Stefanelli, « Cyanotoxins: producing organisms, occurrence, toxicity, mechanism of action and human health toxicological risk evaluation », Archives of Toxicology, vol. 91, no 3, , p. 1049–1130 (ISSN 0340-5761 et 1432-0738, DOI 10.1007/s00204-016-1913-6, lire en ligne, consulté le )
- (en) I. Oksanen, J. Jokela, D. P. Fewer et M. Wahlsten, « Discovery of Rare and Highly Toxic Microcystins from Lichen-Associated Cyanobacterium Nostoc sp. Strain IO-102-I », Applied and Environmental Microbiology, vol. 70, no 10, , p. 5756–5763 (ISSN 0099-2240, PMID 15466511, PMCID PMC522101, DOI 10.1128/AEM.70.10.5756-5763.2004, lire en ligne, consulté le )
- (en) Hans W. Paerl, Timothy G. Otten et Alan R. Joyner, « Moving towards adaptive management of cyanotoxin-impaired water bodies », Microbial Biotechnology, vol. 9, no 5, , p. 641–651 (PMID 27418325, PMCID PMC4993183, DOI 10.1111/1751-7915.12383)
- Stewart, I., Seawright, A. A. et Shaw, G. R. (2008). Cyanobacterial poisoning in livestock, wild mammals and birds–an overview. In Cyanobacterial harmful algal blooms: State of the science and research needs (pp. 613-637). Springer New York.
- Codd GA, Bell SG, Kaya K, Ward CJ, Beattie KA, Metcalf JS (1999), Cyanobacterial toxins, exposure routes and human health. Eur. J. Phycol., 34(4):405–415.
- R Cauchie, HM Hoffmann Les cyanobactéries (algues bleues) des eaux stagnantes du bassin Artois-Picardie et des régions avoisinantes Centre de recherche public Gabriel-Linnmann, Agence de l'eau Artois Picardie, présentation (PDF, 34 pp), consultée 2013-10-03
- Vezie et al. (1997), Occurrence of Microcystin-containing cyanobacterial blooms in freshwaters of Brittany (France), Archiv fur Hydrobiologie 139(3):401-413; janvier 1997
- Brient et al. (2008), A phycocyanin probe as a tool for monitoring cyanobacteria in freshwater bodies ; Journal of Environmental Monitoring (Impact Factor: 2.18). 03/2008; 10(2):248-55, DOI 10.1039/b714238b
- C. Svrcek et D.W. Smith, Cyanobacteria toxins and the current state of knowledge on water treatment options: a review, J. Environ. Eng. Sci., 3: 155-184, 2004.
- Höger SJ (2003), konstanz.de/v13/volltexte/2003/1071//pdf/SJHoeger_Thesis.pdf Problems during drinking water treatment of cyanobacterial loaded surface waters: consequences for human health. Doctoral Thesis Constance : Universität Konstanz
- Falconer I.R. (2005), Cyanobacterial toxins of drinking water supplies: cylindrospermopsins and microcystins, Boca Raton, CRC Press.
- Gorham PR, Carmichael WW (1988), Hazards of freshw ater blue–green algae (cyanobacteria). In: Algae and human affairs. Edited by Lembi CA, Waaland JR. Cambridge: Cambridge University Press 403–431
- Orr PT, Jones GJ, Hunter RA, Berger K, De Paoli DA, Orr CL. (2001), Ingestion of toxic Microcystis aeruginosa by dairy cattle and the implications for microcystin contamination of milk, Toxicon (en). décembre 2001 ; 39(12):1847-54.
- Elliott J (2001), ?file_uuid=2FC47F6C-9D1A-FA75-465E-DE8DCA75BBD4&siteName=aahc Cyanobacterial toxicity. An imal Health Surveillance Quarterly Report 6(1):13
- Duy TN, Lam PKS, Shaw GR, Connell DW (2000), Toxicology and risk assessment of freshwater cyanobacterial (blue-green algal) toxins in water. Rev En- viron Contam Toxicol 163:113–185
- World Health Organisation (WHO), Toxic cyanobacteria in Water: A guide to their public health consequences, monitoring and management, 1re éd., 1999.
- S. Hiller, B. Krock, A.Cembella, B. Luckas, Rapid detection of cyanobacterial toxins in precursor ion mode by liquid chromatography tandem mass spectrometry. Journal of mass spectrometry : 42 (9), 1238-1250, Sep 2007.
- Agence française de sécurité sanitaire de l’environnement et du travail (AFSSET), Évaluation des risques liés à la présence de cyanobactéries et de leurs toxines dans les eaux destinées à l’alimentation, à l’eau de baignade et aux autres activités récréatives, juillet 2006.
- Francis, G. (1878). Poisonous australian lake. Nature, 18, 11-12.
- Codd GA, Lindsay J, Young FM, Morrison LF, Metcalf JS (2005), Harmful cyanobacteria: from mass mortalities to management measures. In: Harmful cyanobacteria. Edited by Huisman J, Ma tthijs HCP, Visser PM. Dordrecht: Springer 1–23
- Braun, A. et Pfeiffer, T. (2002). Cyanobacterial blooms as the cause of a Pleistocene large mammal assemblage. Paleobiology, 28(1), 139-154.
- Cynthia Marshall Faux et Kevin Padian (2007), The opisthotonic posture of vertebrate skeletons: postmortem contraction or death throes ?. Paleobiology 33:2, 201-226 Online publication date: 1-Mar-2007 (résumé)
- Vezie, C., Brient, L., Sivonen, K., Bertru, G., Lefeuvre, J. C. et Salkinoja-Salonen, M. (1997). Occurrence of microcystin-containing cyanobacterial blooms in freshwaters of Brittany (France). Archiv für Hydrobiologie, 139(3), 401-413 (résumé avec Inist/CNRS)
- Briand, J. F., Robillot, C., Quiblier-Lloberas, C. et Bernard, C. (2002), A perennial bloom of Planktothrix agardhii (Cyanobacteria) in a shallow eutrophic French lake : limnological and microcystin production studies. Archiv für Hydrobiologie, 153(4), 605-622 (résumé Inist/CNRS)
- Griffiths DJ, Saker ML (2003) The Palm Island mystery disease 20 years on: a review of research on the cyanotoxin cylindrospermopsin. Environ. Toxicol., 18(2):78–93 (résumé)
- L. A. Lawton, C. Edwards, Review: Purification of microcystins, Journal of Cromatography A 912: 191-209, 2001.
- Codd GA, Edwards C, Beattie KA, Barr WM, Gunn GJ (1992), Fatal attraction to cyanobacteria ? Nature 359(6391):110–111
- Lopez Rodas V, Costas E (1999), Preference of mice to consume Microcystis aeruginosa (toxin–producing cyanobacteria): a possible explanation for numerous fatalities of livestock and wildlife, Res. Vet. Sci., 67(1):107–110 (résumé)
- Corkill N, Smith R, Seckington M, Pontef ract R (1989), Poisoning at Rutland Water. Vet Rec 125(13):356
- Codd GA, Metcalf JS, Morrison LF, Krieni tz L, Ballot A, Pflugmacher S, Wiegand C et Kotut K (2003), Susceptibility of flamingos to cyanobacterial toxins via feeding, Vet. Rec., 152 (23), 722–723.
- Dos S Vieira JM, de P Azevedo MT, de Oliveira Azevedo SM, Honda RY, Corrêa B. (2005), Toxic cyanobacteria and microcystin concentrations in a public water supply reservoir in the Brazilian Amazonia region. Toxicon. 2005 Jun 1; 45(7):901-9. Epub 2005 Apr 14.
- Creasia D.A. (1990), Acute inhalation toxicity of microcystin–LR with mice, Toxicon, 28(6):605.
- Ito E, Kondo F, Harada K (2001), Intratracheal administration of microcystin–LR, and its distribution, Toxicon, 39(2–3):265–271
- Drobac D. et al. (2017), Microcystin accumulation and potential effects on antioxidant capacity of leaves and fruits of Capsicum annuum ; Journal of Toxicology and Environmental Health, Part A Current Issues, vol. 80, no 3 (résumé)
- University of Southern Denmark (2013), Invasive mussel not harmed by toxins, invades freshwaters of Europe, North America, ScienceDaily, 3 octobre 2013. www.sciencedaily.com/releases/2013/10/131003111155.htm
- Michigan State University (2004), Lakes With Zebra Mussels Have Higher Levels Of Toxins, ScienceDaily, 11 mars 2004. www.sciencedaily.com/releases/2004/03/040311072359.htm
- Assessing potential health risks from microcystin toxins in blue-green algae dietary supplements, Environ. Health Perspect., mai 2000, 108(5): 435–439. D J Gilroy, K W Kauffman, R A Hall, X Huang et F S Chu, lire en ligne.
- Contamination by Microcystis and microcystins of blue-green algae food supplements (BGAS) on the italian market and possible risk for the exposed population, Food Chem. Toxicol., décembre 2012, 50(12):4493-9. DOI 10.1016/j.fct.2012.09.029. Epub 2 octobre 2012, Vichi S, Lavorini P, Funari E, Scardala S, Testai E.
- Toxin content and cytotoxicity of algal dietary supplements. Toxicol. Appl. Pharmacol., 2012 Dec 1;265(2):263-71. DOI 10.1016/j.taap.2012.10.005. Epub 2012 Oct 12. Heussner AH, Mazija L, Fastner J, Dietrich DR.
- Codd GA (1983), Cyanobacterial poisoning hazard in British freshwaters. Vet Rec 113(10):223–224
Voir aussi
Articles connexes
Bibliographie
- (en) Codd GA, Lindsay J, Young FM, Morrison LF, Metcalf JS (2005), Harmful cyanobacteria: from mass mortalities to management measures. Dans Harmful cyanobacteria. Edited by Huisman J, Matthijs HCP, Visser PM. Dordrecht : Springer 1–23
- (en) Cook WO, Beasley VR, Lovell RA (1989), Consistent inhibition of peripheral cholinesterases by neurotoxins from the freshwater cyanobacterium Anabaena flosaquae: studies of ducks, swine, mice, and a steer. Environ. Toxicol. Chem., 8(10):915–922
- (en) Cook WO, Iwamoto GA, Schaeffer DJ, Carmichael WW, Beasley VR (1990), Pathophysiologic effects of anatoxin–a(s) in anaesthetized rats: the influence of atropine and artificial respiration. Pharmacol. Toxicol., 67(2):151–155
- (en) Drobac D et al. (2017), Microcystin accumulation and potential effects on antioxidant capacity of leaves and fruits of Capsicum annuum ; Journal of Toxicology and Environmental Health, Part A Current Issues, vol. 80, no 3 (résumé)
- (en) Gugger M, Lenoir S, Berger C, Ledreux A, Druart JC, Humbert JF, Guette C, Bernard C (2005), First report in a river in France of the benthic cyanobacterium Phormidium favosum producing anatoxin–a associated with dog neurotoxicosis. Toxicon, 45(7):919–928
- (en) Hayashi O, Katoh T, Okuwaki Y (1994), Enhancement of antibody production in mice by dietary Spirulina platensis. J Nutr Sci Vitaminol (Tokyo) 40(5):431–441
- (en) Henriksen P, Carmichael WW, An J, Moestrup O (1997), Detection of an anatoxin–a(s)–like anticholinesterase in natural blooms and cultures of cyanobacteria/blue–green algae from Danish lakes and in the stomach contents of poisoned birds. Toxicon, 35(6):901–913