Méristème
Chez les plantes terrestres, un méristème (du grec μεριστός, meristos, « divisé ») est une zone de division cellulaire, à l’origine d’organes et/ou de tissus végétaux (ce n’est pas un tissu car formé de cellules indifférenciées)[1]. Les cellules méristématiques indifférenciées se divisent (mitoses) puis se différencient en acquérant une structure et une fonction. On distingue habituellement les méristèmes primaires qui assurent la croissance de la plante en longueur au niveau de la tige, des feuilles et des racines, et les méristèmes secondaires, responsables de la croissance en épaisseur de certains organes (dits à croissance secondaire) chez certaines plantes (le tronc des arbres par exemple)[2]. Chez les végétaux, les méristèmes permettent une croissance infinie.
Description
Un méristème est un groupe de cellules végétales indifférenciées qui ont la capacité de se diviser par mitose un nombre indéfini de fois[3]. Les cellules du méristème sont aussi appelées "initiales"[4].
Méristèmes et développement
Mousses et ptéridophytes
Les mousses et les ptéridophytes (les fougères entre autres), possèdent une cellule apicale unique de forme tétraédrique présentant donc quatre faces. À chaque cycle cellulaire, cette cellule se divise et donne une nouvelle cellule sur une de ses faces. La nouvelle cellule suivante sera produite sur une autre face, et ainsi de suite ; l'ordre de succession des faces est constant.
Gymnospermes
Les gymnospermes (les conifères entre autres) possèdent des méristèmes constitués de deux couches de cellules : une première couche en surface dont les cellules subissent des divisions anticlines uniquement (l'axe de la séparation des cellules est perpendiculaire à la surface du méristème), et une couche sous-jacente dont les divisions sont anticlines et périclines (l'axe de la séparation des cellules est parallèle à la surface du méristème).
Angiospermes
Les plantes à fleurs possèdent des méristèmes similaires à ceux des gymnospermes, mais avec une couche intermédiaire (L2) entre la couche externe (L1) et la couche interne (L3) dont les cellules ont une division majoritairement anticline. (voir plus loin l'organisation des tissus méristématiques)
La biologie du développement chez les plantes (en) montre différents stades de développement : d'abord les méristèmes floraux se différencient du méristème d'inflorescence puis se mettent en place les primordia des organes floraux.
Histogenèse et organogenèse
Les méristèmes forment des tissus (histogenèse) tels que des tissus conducteurs (xylème, phloème), et parfois des organes entiers (organogenèse) comme les feuilles, eux-mêmes constitués de tissus..
Les méristèmes (primaires comme secondaires) produisent de nouvelles cellules continuellement, en dehors de leurs périodes de quiescence qui se traduisent par une interruption de leur activité.
La formation des feuilles débute avec la mise en place au niveau des méristèmes caulinaires (aériens) d'ébauches foliaires, des massifs de cellules programmés pour évoluer en feuilles.
Les cellules créées par les méristèmes (on appelle ce processus la mérèse) se différencieront progressivement, notamment par le biais de l'auxèse.
Chaque méristème peut potentiellement croître indéfiniment ; ce qui implique que la forme complète de la plante n'est pas déterminée à l'avance. Elle peut ainsi s'ajuster aux conditions environnantes, contrairement aux animaux bilatériens qui disposent d'un plan d'organisation cellulaire défini. Ils sont aussi responsables de la disposition particulière des feuilles et branches autour de la tige (phyllotaxie).
Les différents méristèmes
On peut distinguer de manière générale :
- Les méristèmes primaires qui sont organogènes et histogènes
- Les méristèmes secondaires qui sont seulement histogènes.
Méristèmes primaires ou apicaux
Les méristèmes primaires sont les premiers à se mettre en place ; ils sont à l'origine des tissus primaires de la plante et permettent la croissance en longueur. Ils sont situés à l'apex des racines et de la tige et des rameaux (dans les bourgeons lorsqu'il y en a - ils sont alors quiescents). On les trouve aussi dans les bourgeons axillaires, à l'aisselle des feuilles.
Méristèmes secondaires ou latéraux
Ils ne sont pas présents chez tous les végétaux ; les plantes herbacées par exemple n'ont pas de croissance secondaire.
Le méristème secondaire n'existe que chez les gymnospermes et les angiospermes dicotylédones
Les méristèmes secondaires (cambium et phellogène) sont mis en place après que la croissance en longueur est achevée dans la zone. Ils vont alors déclencher la croissance en épaisseur de la plante.
Dans les tiges des dicotylédones, le cambium provient d'une part de cellules restées procambiales placées entre le xylème primaire et le phloème, et d'autre part de la dédifférenciation de cellules parenchymateuses qui se dédifférencient entre les massifs de tissus conducteur pour "fermer le cercle du cambium"[5].
Dans les racines des dicotylédones, le cambium provient d'une part de cellules restées procambiales placées en face des massifs de phloème primaire, et d'autre part de la prolifération et de la différenciation de certaines cellules du péricycle en face des massifs de xylème primaires. Il a alors une forme étoilé, et deviendra rapidement circulaire[6].
Les cellules qui le constituent subissent des divisions périclines, de manière à créer :
- vers l'intérieur : du xylème secondaire, lignifié, tissu conducteur de la sève brute et de soutien contenant notamment des vaisseaux et des fibres.
- vers l'extérieur : du phloème secondaire (liber), tissu conducteur de la sève élaborée et de soutien.
Les vaisseaux primaires sont comprimés par la croissance des tissus secondaires.

De nouveaux vaisseaux sont ainsi produits, remplaçant les précédents qui disparaissent dans le cas du phloème et restent en place bien que dysfonctionnels dans le cas du xylème, participant au soutien de la plante. Ces anciens vaisseaux forment le bois de cœur (duramen). Cette croissance varie selon les conditions environnementales, conduisant à des anneaux plus ou moins foncés selon l'intensité de l'activité de la zone cambiale dans les régions tempérées.
Les anneaux foncés sont créés lors des périodes de faible croissance, et sont attribuables à un manque de ressource pour le cambium, comme le manque d'eau ou le passage d'un hiver. Les anneaux plus clairs, créés lors de périodes de forte croissance, correspondent à des vaisseaux d'épaisseur plus importante engendrés lorsque le cambium dispose de ressources plus importantes.
L'alternance de périodes favorables (printemps, été) et défavorables (automne, hiver) est à l'origine de la formation de deux types de bois :
- Le bois de printemps, constitué en début de saison de végétation, en général de couleur claire ;
- Le bois d'été, constitué en fin de saison de végétation, de couleur plus sombre. C'est la superposition de ces deux couches de bois qui forme le cerne.
Le décompte des cernes d'un arbre peut ainsi donner une idée particulièrement précise de son âge: c'est ce principe qui est utilisé en dendrochronologie (figure 1).
Le deuxième méristème secondaire est le phellogène ou assise génératrice subéro-phellodermique. Il se forme dans la tige à partir de la dédifférenciation de cellules du collenchyme et du parenchyme cortical (de l'écorce) placées sous l'épiderme ; et dans la racine à partir de la dédifférenciation du péricycle[7].
Celui-ci, comme le cambium, est le siège de divisions périclines, créant :
- vers l'intérieur : du phelloderme ayant pour fonction la régénération du phellogène.
- vers l'extérieur : du suber, (liège) ayant un rôle de protection de la plante, qui s'ajoute à celle que procure l'épiderme.
Organisation du tissu méristématique
Cellules
Les cellules méristématiques ont des fonctions analogues à celles des cellules souches chez les animaux : elles sont peu ou pas du tout différenciées et sont capables de se diviser indéfiniment par division cellulaire . La différence existant entre les cellules souches et les cellules spécialisées, qui n'ont plus le potentiel de différenciation des cellules souches, est beaucoup moins marquée chez les plantes : de nombreuses cellules différenciées des plantes sont capables de se dédifférencier et de reconstruire de nouveaux tissus ou organes. Le bouturage est un exemple de ces capacités.
Ces cellules sont indifférenciées, ce qui se traduit notamment par des vacuoles très réduites et une absence de plastes différenciés (ils sont présents sous une forme rudimentaire). Les cellules méristématiques sont étroitement accolées les unes aux autres, séparées par des parois pectocellulosiques très fines et ne présentant aucune cavité intercellulaire. Leur rapport nucléocytoplasmique (rapport des surfaces du noyau et du cytoplasme en coupe) est de l'ordre de 1[8].
Structure du méristème
Le méristème apical caulinaire peut être divisé en couches ou en domaines :
Couches
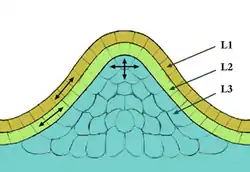
L'organisation en couches prend en compte les différentes lignées cellulaires qui constituent chacune une couche(figure 2).
La couche de cellules la plus externe (couche L1) constitue l'épiderme et la couche sous-jacente est la couche L2. La couche la plus interne, formée de plusieurs couches de cellules, constitue la couche L3, qui engendrera le parenchyme médullaire. Les couches L1 et L2 subissent toutes deux des divisions anticlines et forment la tunica du méristème, tandis que la couche L3 qui présente des divisions à la fois anticlines et périclines constitue le corpus du méristème.
Domaines

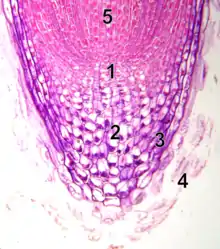
1 - zone centrale
2 - zone périphérique
3 - méristème médullaire
4 - tissus médullaires
Une autre présentation du méristème consiste à le diviser en domaines (ou zones) selon la fonction des cellules. Certains considèrent ainsi que le méristème apical est plutôt un ensemble de méristèmes spécialisés dans la formation d'un type de cellule spécifique (figure 3).
Les cellules de la zone centrale (ZC) (1) (ou centre quiescent) possèdent une faible activité mitotique et forment des cellules totalement indifférenciées. Celles-ci se divisent et migrent vers la zone périphérique (ZP) du méristème (2). Les cellules de cette zone périphérique sont plus actives et forment des primordia foliaires qui donneront les feuilles (rôle organogène du méristème), ainsi que les cellules qui donneront les tissus de l'écorce et les tissus conducteurs[8]. Sous la zone centrale se trouve le méristème médullaire (3), qui assure la création du parenchyme médullaire [8](4). Son rôle est donc histogène.
Régulation de l'activité du méristème
L'activité du méristème est régulée de manière à ne pas épuiser ses ressources ou produire trop de cellules. En outre, les différents méristèmes ont une identité déterminée (MAC, MAR, méristème floral…). Tout cela est réalisable à l'aide de signaux.
Biotechnologies
Le méristème est un tissu important en biotechnologie. En effet, comme on ne trouve pas de virus dans les méristèmes d'une plante infectée, on peut régénérer une plante saine à partir d'une plante malade en faisant une culture in vitro de son méristème. La culture de méristèmes pour la canne à sucre, le manioc, l'igname ou le bananier est ainsi indispensable pour maintenir des populations saines dans les zones tropicales, particulièrement exposées aux épiphyties (épidémies végétales)[9].
Ce procédé est en outre très utilisé en horticulture pour produire des plantes d'un même génotype. C'est une technique de multiplication asexuée, ou végétative.
Notes et références
- Jacques Margara, Bases de la multiplication végétative : les méristèmes et l'organogenèse, Paris, INRA, , 262 p. (ISBN 2-85340-379-3)
- Morphologie végétale : méristèmes UC Louvain
- Pascal Gantet et Alain Verger, « Méristèmes »
 , sur Unisciel, (consulté le )
, sur Unisciel, (consulté le ) - Integrative Plant Anatomy, page 3
- Botanique, biologie et physiologie végétales p.81 (2e édition, 2008) S.MEYER, C.REEB, R.BOSDEVEIX
- Botanique, biologie et physiologie végétales p.71 (2e édition, 2008) S.MEYER, C.REEB, R.BOSDEVEIX
- Botanique, biologie et physiologie végétales p.71 et 82 (2e édition, 2008) S.MEYER, C.REEB, R.BOSDEVEIX
- Dictionnaire raisonné de biologie (2002) , MORERE J.-L. , PUJOL R
- P. Mollier, Les biotechnologies vertes, nouvelles pistes pour l’agriculture, site de l'INRA, le 8 février 2013