Hemilepistus reaumuri
Cloporte de Réaumur

| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Sous-embr. | Crustacea |
| Classe | Malacostraca |
| Ordre | Isopoda |
| Famille | Agnaridae |
| Genre | Hemilepistus |
- Porcellio reaumuri H. Milne-Edwards, 1840
(protonyme) - Hemilepistus bodenheimeri
Verhoeff, 1931 - Hemilepistus palaestinus Verhoeff, 1931
- Paraniambia tuberculata Collinge, 1914
Hemilepistus reaumuri, parfois appelé cloporte de Réaumur, est une espèce de cloportes de la famille des Agnaridae, vivant à l'intérieur et aux alentours des déserts d'Afrique du Nord et du Moyen-Orient, soit l'habitat le plus sec conquis par toute espèce de crustacé. Il atteint une longueur de 22 mm pour une largeur maximale de 12 mm, tandis que ses sept paires de pattes maintiennent son corps particulièrement surélevé du sol. L'espèce apparaît dans la Description de l'Égypte, ouvrage rassemblant des découvertes effectuées par la Commission des sciences et des arts lors de l'expédition d'Égypte de 1798 à 1801, mais n'a été décrite formellement qu'en 1840, par Henri Milne Edwards, qui lui a donné le nom de Porcellio reaumuri. Il a pris son nom scientifique actuel quand l'ancien sous-genre Hemilepistus a été élevé au rang de genre en 1930.
H. reaumuri forme des populations très denses et occupe une niche écologique importante dans l'écosystème désertique. Il se nourrit de feuilles, assouvit en grande partie ses besoins en eau par l'humidité de l'air et du sable et représente une proie importante pour le scorpion Scorpio maurus. Ce sont les soins parentaux accordés à la progéniture qui permettent à H. reaumuri de survivre dans des conditions extrêmement arides. Les adultes creusent des terriers où vivent des groupes familiaux, socialement soudés par l'usage de phéromones. Les nids atteignent 40 à 50 cm de profondeur et les cloportes profitent des conditions relativement fraîches et humides du terrier lorsque les conditions de surface sont défavorables. Les limites du territoire de chaque colonie sont indiquées par des tas de matières fécales.
Description
Hemilepistus reaumuri mesure environ 22 millimètres de long et 9 à 12 millimètres de large[1]. Comme tous les isopodes, il compte sept paires de pattes et une paire d'antennes bien visibles[2]. Il a des poumons à l'intérieur des cinq paires d'appendices abdominaux, ce qui lui vaut d'être placé dans la famille des Agnaridae. Il est placé dans le genre Hemilepistus en raison de la présence de tubercules seulement sur la tête et sur les parties voisines du thorax[3].
H. reaumuri diffère des autres cloportes du désert à un certain nombre d'égards. Il est crépusculaire, tandis que les autres sont nocturnes[4]. En dehors des températures les plus élevées, il est également photophile (il est attiré par la lumière du soleil), alors que les autres espèces fuient la lumière vive[4]. De plus, comparé à d'autres cloportes, H. reaumuri est très haut sur pattes[5].
Écologie et comportement
Mode de vie

Hemilepistus reaumuri ne peut échapper à la chaleur du désert qu'en construisant un terrier, ce qui est coûteux en temps et en énergie. Un parent garde donc le nid pendant que l'autre part à la recherche de nourriture[6]. Ce cloporte peut passer jusqu'à près de dix mois de l'année à revenir à la surface pour se nourrir, alors que les espèces qui ne creusent pas de terriers, comme Armadillidium vulgare ou Armadillo officinalis, ont des périodes d'activité à l'extérieur plus courtes[4]. Bien qu'ils prospectent jusqu'à des températures de 35 °C, ils retournent à leur terrier quand la température est trop élevée[4]. Ils sont également incapables de tolérer l'air avec une humidité relative inférieure à 6 %, ce qui se produit souvent à des profondeurs allant jusqu'à 30 centimètres dans le sol du désert au cours des mois les plus chauds : les nids font donc au moins 40 à 50 cm de profondeur[4]. Les terriers sont verticaux, avec une seule entrée de 9 à 12 mm de diamètre[7], et il peut y avoir jusqu'à 20 terriers par mètre carré dans les zones favorables[8]. Si un cloporte a du mal à trouver l'entrée de son nid à son retour, il emploie une stratégie complexe et efficace pour le localiser. Il commence par décrire une spirale s'élargissant autour de l'endroit où le terrier devrait se trouver et, plus il met de temps, plus il adopte une trajectoire sinueuse[7].
Ces cloportes passent leurs journées à ramener à leur terrier des feuilles trouvées en surface dans le désert, restant parfois au repos sous des pierres ou dans des anfractuosités de rochers[4]. Leurs déjections s'accumulent à la surface et forment un cercle de matière fécale autour du trou, semblable à une digue, qui délimite l'étendue du territoire de la colonie du terrier[8]. L'armoise herbe blanche (Artemisia herba-alba) et la saligne à balai (Hammada scoparia) sont les buissons les plus abondants dans le désert du Néguev et semblent constituer l'essentiel du régime alimentaire de H. reaumuri[9].
Hemilepistus reaumuri trouve essentiellement l'eau dont il a besoin dans la vapeur d'eau de l'air saturé et en mangeant du sable humide. La perte d'eau est minimisée par l'épithélium rectal, qui absorbe l'eau, assurant que les fèces soient plus sèches que la nourriture consommée[10]. L'évaporation de l'eau à travers l'exosquelette perméable pourrait toutefois fournir un effet de refroidissement précieux[5].
Niche écologique

Hemilepistus reaumuri représente une partie importante de l'écosystème du désert. Les densités de population peuvent atteindre 480 000 individus par hectare, ce qui équivaut à une biomasse de 19,2 kg/ha ; à titre de comparaison, l'ensemble des mammifères du désert représente selon les estimations une biomasse totale de 39,9 kg/ha[11]. Ce cloporte a une biomasse significativement plus élevée que les autres herbivores dans le désert du Néguev, ce qui en fait un maillon important de la chaîne alimentaire herbivores-omnivores[4].
Le principal prédateur de H. reaumuri est le scorpion Scorpio maurus[12], dont le cloporte peut représenter jusqu'à 70 % de l'alimentation[13]. Ce crustacé ne semble être vulnérable aux attaques qu'en surface : on ne connaît pas de prédateurs attaquant H. reaumuri dans son terrier[12].
Reproduction
Hemilepistus reaumuri est la seule espèce de son genre à présenter des soins parentaux pour la progéniture[14]. Le développement de ce trait a été crucial pour que l'espèce puisse survivre dans le désert[6]. Ce cloporte est monogame et, chose exceptionnelle, les deux parents veillent au soin des jeunes[6].
Les colonies sont au repos pendant l'hiver et les jeunes individus sortent en février et mars pour établir de nouveaux terriers[8]. Ils choisissent des sites abrités par des buissons ou certains grands mâles tentent de se rapprocher d'une femelle ayant déjà établi un abri, évinçant parfois le partenaire si celle-ci en a un[8]. L'anatomie de H. reaumuri n'est pas adaptée au fouissage et l'excavation est un processus lent, qui a lieu seulement au début du printemps[15]. Les 3 à 5 premiers centimètres sont creusés par un cloporte seul, qui s'arrête alors pour garder le nouveau terrier. Il finit par laisser un individu du sexe opposé entrer et tous deux s'engagent alors dans un rituel qui dure souvent pendant des heures, débouchant sur la copulation[15].
La femelle donne naissance à 50 à 100 jeunes, généralement en mai. Ceux-ci restent dans le terrier pendant 10 à 20 jours, nourris par les parents. En quittant le nid, ils se méfient des autres familles, les évitant dans un premier temps ; les adultes peuvent attraper les jeunes d'autres terriers pour les manger ou en nourrir leur propre progéniture, mais n'attaquent pas cette dernière[15]. En effet les membres de chaque groupe social se reconnaissent mutuellement en utilisant des phéromones[4]. Chaque couple ne donne naissance qu'à une portée et la durée de vie d'un individu est généralement proche de 15 mois, ce qui est considérablement plus court que les 2 à 4 ans qu'atteignent les cloportes d'habitats mésiques (modérément humide), comme Armadillidium vulgare, Porcellio scaber ou Philoscia muscorum[16].
Répartition et habitat

Cette espèce vit dans les steppes, les semi-déserts et les déserts d'Afrique du Nord et du Moyen-Orient, occasionnellement sur les bords des lacs salés[15], soit « l'habitat le plus sec conquis par toute espèce de crustacé »[14]. H. reaumuri est le plus étroitement associé à des sols de lœss du Sahara et du Néguev[4], bien que son aire de répartition s'étende de l'Est de l'Algérie à l'Ouest de la Syrie[17].
Taxinomie
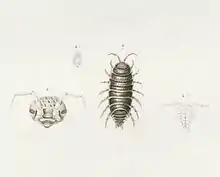
Hemilepistus reaumuri a d'abord été illustré dans le volume 21 de la Description de l'Égypte, série d'ouvrages rapportant les recherches effectuées lors de la campagne d'Égypte menée par Napoléon de 1798 à 1801. La partie sur les crustacés est commencée par Jules-César Savigny, mais la santé de celui-ci se détériorant, c'est Victor Audouin qui termine le livre. Il y mentionne le cloporte sans lui donner de nom scientifique ni faire de diagnose, mais en décidant de lui donner le nom de René-Antoine Ferchault de Réaumur, naturaliste français[18].
C'est finalement Henri Milne Edwards qui décrit formellement l'espèce en 1840, en lui donnant le protonyme de Porcellio reaumuri et en l'appelant « Porcellion de Réaumur »[17] - [19]. Du genre Porcellio, l'espèce est ensuite déplacée par G.H.A. Budde-Lund en 1879 dans un sous-genre nouveau, Hemilepistus[17], élevé au rang de genre par Karl Wilhelm Verhoeff en 1930[20]. Le genre est lui-même initialement placé parmi les Trachelipodidae, puis chez les Porcellionidae à partir de 1989, mais une étude de 2003 revoit toute la systématique des cloportes et donne le jour à la famille des Agnaridae où Hemilepistus est placé[21] - [17].
Plusieurs noms auparavant considérés comme synonymes de H. reaumuri ont vu leur matériel ré-examiné en 2011 et finalement attribué à une espèce d'une autre famille, Porcellio brevicaudatus[3].
Annexes
Bibliographie
- (en) K. Eduard Linsenmair, « Some adaptations of the desert woodlouse Hemilepistus reaumuri (Isopoda, Oniscoidea) to desert environment », Verhandlungen der Gesellschaft für Ökologie, vol. 4, , p. 183–185 (lire en ligne)
- (en) Moshe Shachak, E. A. Chapman et Y. Steinberger, « Feeding, energy flow and soil turnover in the desert isopod, Hemilepistus reaumuri », Oecologia, vol. 24, no 1, , p. 57–69 (DOI 10.1007/BF00545487, JSTOR 4215268)
- (en) Z. Dubinsky, Y. Steinberger et M. Shachak, « The survival of the desert isopod Hemilepistus reaumurii (Audouin) in relation to temperature (Isopoda, Oniscoidea) », Crustaceana, vol. 36, no 2, , p. 147–154 (DOI 10.1163/156854079X00339, JSTOR 20103376)
- (en) Moshe Shachak, « Energy allocation and life history strategy of the desert isopod H. reaumuri », Oecologia, vol. 45, no 3, , p. 404–413 (DOI 10.1007/BF00540214, JSTOR 4216114)
- (en) Gerhard Hoffmann, « The random elements in the systematic search behavior of the desert isopod Hemilepistus reaumuri », Behavioral Ecology and Sociobiology, vol. 13, no 2, , p. 81–92 (DOI 10.1007/BF00293798, JSTOR 4599612)
- (en) Gerhard Hoffmann, « The search behavior of the desert isopod Hemilepistus reaumuri as compared with a systematic search », Behavioral Ecology and Sociobiology, vol. 13, no 2, , p. 93–106 (DOI 10.1007/BF00293799, JSTOR 4599613)
- (en) L.B. Holthuis, « Hemilepistus reaumurii (H. Milne Edwards, 1840), its author(s) and its nomenclatural status (Isopoda terrestria) », Crustaceana, vol. 78, no 10, , p. 1269-1271 (DOI 10.1163/156854005775903609)
- (en) Ghasem M. Kashani, Johann-Wolfgang Wägele et Helmut Schmalfuss, « Redescription of Porcellio brevicaudatus Brandt, 1833 (Isopoda: Oniscidea); with some notes on other synonyms of Hemilepistus reaumurii (Milne-Edwards, 1840) », Zootaxa, vol. 2924, , p. 63–67
Références taxinomiques
- (en) Référence Animal Diversity Web : Hemilepistus reaumuri
- (en) Référence Catalogue of Life : Hemilepistus reaumuri (Milne Edwards, 1840) (consulté le )
- (fr+en) Référence ITIS : Hemilepistus reaumuri (Audouin, 1826)
- (en) Référence World Register of Marine Species : espèce Hemilepistus reaumuri (Milne Edwards, 1840))
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Hemilepistus reaumuri » (voir la liste des auteurs).
Notes
- L'auteur est souvent cité, à tort, comme « (Audouin, 1826) » ou « (Audouin & Savigny, 1826) », voir la section Taxinomie pour plus d'explications.
Références
- Hoffmann (1983 a)
- (en) Maurice Burton et Robert Burton, The International Wildlife Encyclopedia, vol. 1, Marshall Cavendish, (ISBN 978-0-7614-7266-7, lire en ligne), « Wood louse », p. 2973–2973
- Kashani, Wägele et Schmalfuss (2011)
- (en) Fred Punzo, Desert Arthropods : Life History Variations, Springer, (ISBN 978-3-540-66041-5, lire en ligne), « Life histories: individual case studies », p. 105–156
- (en) E. B. Edney, « Animals of the desert », dans Edwin Sherbon Hills, Arid Lands: a Geographical Appraisal, Taylor & Francis, , 461 p. (ISBN 0-416-28650-X, lire en ligne), p. 181–218
- (en) Tristram Wyatt, « How habitat leads to mothercare », New Scientist, vol. 116, no 1581, , p. 50–53 (lire en ligne)
- Hoffmann (1983 b)
- (en) James T. Costa, The Other Insect Societies, Harvard University Press, , 767 p. (ISBN 978-0-674-02163-1, lire en ligne), « Other social arthropods. Arachnids, centipedes, millipedes, and crustaceans », p. 667–716
- Shachak, Chapman et Steinberger (1976)
- (en) Colin Little, The Colonisation of Land : Origins and Adaptation of Terrestrial Animals, Cambridge University Press, , 290 p. (ISBN 978-0-521-25218-8, lire en ligne), « Crustaceans and the evolution of the arthropods », p. 63–106
- (en) James T. Markwiese (dir.), Randall T. Ryti, Mark M. Hooten, Daniel I. Michael et Ihor Hlohowskyj, « Toxicity bioassays for ecological risk assessment in arid and semiarid ecosystems », Reviews of Environmental Contamination and Toxicology, Springer, vol. 168, , p. 43–98 (ISBN 9780387951386, lire en ligne)
- Dubinsky, Steinberger et Shachak (1979)
- (en) David Ward, The Biology of Deserts, Oxford, Oxford University Press, , 339 p. (ISBN 978-0-19-921147-0, lire en ligne), « The importance of predation and parasitism », p. 124–144
- (en) Rod Preston-Mafham et Ken Preston-Mafham, The Encyclopedia of Land Invertebrate Behavior, MIT Press, , 320 p. (ISBN 978-0-262-16137-4, lire en ligne), « Crustacea. Woodlice, crabs », p. 161
- Linsenmair (1974)
- Shachak (1980)
- (en) Helmut Schmalfuss, « World catalog of terrestrial isopods (Isopoda: Oniscidea) – revised and updated version », Stuttgarter Beiträge zur Naturkunde, a, vol. 654, , p. 1–341 (lire en ligne)
- (fr) Victor Audouin, Description de l'Égypte ou recueil des observations et des recherches qui ont été faites en Égypte pendant l'expédition de l'armée française, t. I - Histoire naturelle : L'explication sommaire de planches de Crustacés de l'Égypte et de la Syrie, publiée par Jules-César Savigny, membre de l'Institut, offrant un exposé des caractères naturels des genres, avec la distinction des espèces, Imprimerie impériale (lire en ligne), p. 98
- (fr) Henri Milne Edwards, Histoire naturelle des crustacés, t. 3, Paris, (lire en ligne), p. 170-171
- (en) Ghasem M. Kashani, Alireza Sari et Shidokht Hosseinie, « Terrestrial isopods of the subgenus Hemilepistus (Hemilepistus) Budde-Lund, 1879 (Isopoda: Oniscidea) from Iran », Zootaxa, vol. 2549, , p. 54–68 (lire en ligne)
- (en) Christian Schmidt, « Phylogeny of the terrestrial Isopoda (Oniscidea): a review », Arthropod Systematics & Phylogeny, vol. 66, no 2, , p. 191–226 (lire en ligne [PDF])