Continuum fluvial
Le concept de continuum fluvial (River Continuum Concept), introduit dans les années 1980 par Robin L. Vannote, est un modèle descriptif longitudinal du fonctionnement écologique des cours d’eau des zones tempérées, qui intègre à la fois les indicateurs biologiques et la géomorphologie fluviale. On lui adjoint parfois un continuum sous-fluvial[1].
Le continuum piscicole est la partie du continuum fluvial accessible aux poissons et permettant leur circulation de l'amont à l'aval et réciproquement. Il ne doit pas être confondu avec un continuum physique ou avec la notion ambiguë de libre circulation (tout cours d'eau sauvage étant naturellement hétérogène). Cette libre circulation peut néanmoins être évaluée, par exemple dans les cours d'eau où des réempoissonnements sont fréquents et où l'on pense que des translocations ne sont pas réalisées par des pêcheurs indépendants. On peut y mesurer par des techniques de captures-recaptures, et éventuellement par le suivi de poissons radio-marqués (par exemple expérimenté en Région wallonne dans le cadre de l'inventaire des « obstacles à la libre circulation » des poissons, entamé en 1997 (Ce suivi a confirmé que (hormis l'anguille qui est capable de sortir de l'eau et moindrement les lamproies), la truite franchit le mieux les obstacles, devant l’ombre)[2]. Les lamproies peuvent aussi se "coller" au moyen de leur ventouse à des poissons beaucoup plus grands et plus forts qu'elles pour se faire remonter vers la source[3].
Contexte écopaysager
Le concept de continuum fluvial repose sur une vision holistique qui est que les cours d’eau, de leurs source aux estuaires doivent être considérés comme des écosystèmes ouverts, en interaction avec la mer, la nappe, les berges et leurs ripisylves et tout le contexte éco-hydraulique qui en dépend. Les modifications progressives de leurs conditions physiques (largeur, profondeur, débit, courant, température...) de l’amont à l’aval engendrent un gradient continu des ressources trophiques disponibles qui détermine la distribution des invertébrés, en fonction des différentes stratégies développées par les espèces pour exploiter ces ressources alimentaires.
CM Pringle invite à ne pas uniquement considérer les flux "amont → aval", mais aussi les flux existant de l'aval vers l'amont[4] (avec par exemple des oligoéléments essentiels remontés par les salmonidés par exemple tel que l'iode) et les effets ou cascades d'effets différés d'actions de l'aval sur les écosystèmes de l'amont (extraction de gravier, barrage, introduction volontaire ou non d'espèces invasive, réchauffement de l'eau, introduction de pathogènes ou de perturbateurs endocriniens, pompage ou irrigation causant un rabattement de nappe et par suite un abaissement du niveau des sources... par exemple).
Selon Pringle, omettre cette relation aval vers amont serait « d'autant plus préoccupant que la plupart des rivières sont altérés par les humains dans leurs bassins inférieurs et que les enquêtes écologiques les plus publiés sur les systèmes lotiques concernent les cours d'eau de l'amont »[4] dont on oublie souvent qu'ils peuvent déjà être altéré par des phénomènes biophysiques épars ayant eu lieu ou ayant lieu sur l'aval du cours d'eau (voire en mer). La suppression de barrages de castors a ainsi des conséquences locales, mais aussi sur l'aval et l'amont. « le recyclage des nutriments, la productivité primaire, les tendances régionales de la biodiversité » dans l'amont du bassin versant sont des paramètres qui peuvent être positivement ou négativement affectés par des actions de renaturation ou au contraire délétères en aval du cours d'eau[4].
Des obstacles tels que grands barrages et zones de forte pollution (dont pollution thermique ou pollution lumineuse) sont alors étudiés comme facteurs de fragmentation écologique de ce continuum.
Le Continumm du fleuve physique visible au sol interagit en particulier avec un autre continuum invisible, celui du « compartiment sous-fluvial »[1] occupé par une faune spécifique (site stygofaune) et parfois lui-même considéré comme un corridor biologique à part entière, invisible mais fonctionnellement relié au fleuve dans la plaine alluviale et son lit majeur, interagissant avec la plupart des zones humides (hors nappes-perchées, tourbières de pente ou zones humides perchées formées sur des lits imperméables dont l'alimentation en eau et indépendante du fleuve).
Division fonctionnelle des cours d’eau


Suivant le concept classique et hydro-géomorphologique de continuum fluvial, on peut toujours diviser les systèmes lotiques en trois domaines majeurs : le ruisseau ou torrent puis les eaux courantes de moyenne vallée et enfin les grands fleuves et rivières.
- Sur le domaine amont, le cours d’eau est étroit. Hormis en zones très froide, de très haute montagne et désertiques ou urbaine, sa ripisylve est généralement très développée. Une faible luminosité et de forts taux de dilution des éléments inorganiques peuvent alors inhiber la production de matière organique par photosynthèse (production primaire) mais de grandes quantités de matière organique allochtone tombent dans le ruisseau. Par conséquent, la respiration (transformation de la matière organique en éléments minéraux) est plus importante que la production primaire (rapport Production/Respiration < 1, hétérotrophie). C’est dans ce domaine que la diversité des ressources trophiques disponibles est la plus grande.
La présence d'une espèce-ingénieur le castor (autrefois commun dans une grande partie de l'hémisphère nord tempéré) peut localement ouvrir le milieu à la lumière et remonter le niveau de l'eau, en modifiant le courant et la structure du cours d'eau (segmentation et système en "escalier") ; - Sur le domaine intermédiaire, l’élargissement de la rivière et la diminution des taux de dilution permet à la production de matériel organique autochtone, notamment sous formes d’algues, d’augmenter sensiblement. On a alors un rapport P/R > 1 (autotrophie) ;
- Sur le domaine aval, le matériel organique particulaire est abondant. L’approfondissement du lit et la turbidité limitent la luminosité et la photosynthèse n’a plus lieu qu’à la surface des eaux. Il y a par conséquent retour à l’hétérotrophie.
Distribution des populations invertébrées
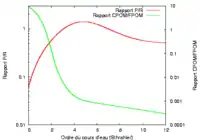
La composition de la biocénose et, plus particulièrement des populations invertébrées, dépend de la position de son biotope le long du continuum fluvial, que l’on mesure par l’ordre du cours d’eau dans la classification de Strahler.
On classe les invertébrés en quatre grandes catégories : les broyeurs, les collecteurs, les brouteurs et les prédateurs. À l’exception des prédateurs, tous ces organismes se nourrissent de matière organique d’origine végétale : ce sont des saprophytes.

Les broyeurs ont pour rôle de déchiqueter le matériel organique grossier (CPOM = Coarse Particulate Organic Matter), de taille millimétrique, tel que des lambeaux de feuillage. Les amphipodes, les aselles, les larves d’éphéméroptères et de plécoptères comptent parmi les broyeurs les plus courants de cours d’eau d’Europe Centrale.
Les collecteurs sont, eux, capables de récolter la matière organique en suspension dans l’eau courante ou piégée dans les sédiments. Les particules dont ils se nourrissent ont une taille comprise entre 0,5 et 50 micrométres (UPOM = Ultrafine Particulate Organic Matter et FPOM = Fine Particulate Organic Matter). Les larves de simulie, les nématodes et de nombreuses larves de chironomidés appartiennent par exemple au groupe des collecteurs.
.jpg.webp)
Les brouteurs se nourrissent de ce qui se développe (algues, micro-organismes…) sur les pierres (le périphyton), les morceaux de bois mort et les grandes plantes aquatiques qui structurent le lit des cours d’eau. Parmi eux, on dénombre les gastéropodes, les larves de liponeura et certaines de chironomidés.
Quant aux prédateurs, comme les larves de libellules et différents coléoptères, ils se nourrissent des autres animaux.
Du fait de l’évolution du matériel organique disponible, la représentation de ces différents groupes varient de l’amont vers l’aval du cours d’eau. En amont, c’est-à-dire dans la partie torrentielle, où le matériel organique grossier est abondant, on trouve une forte proportion de broyeurs mais aussi de collecteurs et les brouteurs sont peu représentés. La proportion de brouteurs augmente ensuite, ainsi que celle des collecteurs, au détriment des broyeurs dont les populations régressent significativement, pour finalement disparaître complètement. En aval, où subsiste presque uniquement le matériel organique particulaire, les collecteurs constituent le groupe dominant. Quant aux prédateurs, leur proportion reste toujours à peu près constante ; seule varie la représentation des espèces au sein du groupe, qui suit l’évolution des proies disponibles. Des variations atypiques dans la répartition des différents groupes invertébrés le long d’un cours d’eau sont le signe d’une perturbation du milieu naturel.

Stabilité du système lotique
Le modèle du continuum fluvial permet de mieux comprendre l’allocation des ressources énergétique dans les systèmes lotiques.
À chaque niveau du cours d’eau, l’énergie disponible sous forme de matériel organique peut être consommée, stockée ou transférée en aval. Comme elle constitue le facteur limitant du système, celui-ci doit assurer son utilisation optimale. Des ressources inutilisées permettent en effet à de nouvelles espèces d’intégrer l’écosystème, jusqu’à saturation de son potentiel énergétique. Ce principe est valable pour tout écosystème de manière générale mais il joue ici un rôle particulier en raison de l’évacuation continue du matériel organique non consommé, emporté par les eaux courantes, qui implique un impératif d’efficacité plus important et surtout une utilisation continue des ressources.
Le problème de la continuité temporelle se pose essentiellement aux échelles de la journée et de l’année. Au cours d’une journée, les risques accrus de prédation (les poissons chassent essentiellement grâce à leur vision) et les modifications de facteurs abiotiques tels que la température et la luminosité, induisent une alternance des espèces actives. Les variations journalières étant plus importante sur les cours d’eau intermédiaires, c’est là que la diversité biologique est la plus importante.
L’optimisation constante de l’utilisation des ressources énergétique dans les systèmes lotiques garantit une grande stabilité vis-à-vis des fluctuations et perturbations de leurs conditions physiques. Après une perturbation, ces systèmes trouvent très rapidement un nouvel équilibre sans succession écologique.
Modélisation de la continuité
Le concept de continuum fluvial a été l'objet de plusieurs années de recherches financées par la fondation Rockefeller et publiées en 1980 sous le titre The River Continuum Concept dans le Canadian Journal of Fisheries and Aquatic Sciences. Un modèle s’est appuyé sur les travaux de Ruth Patrick et de Luna Leopold. L’apport essentiel de l’équipe de Vannote aura été une prise en compte globale et exhaustive de l’écosystème lotique.
Après leur publication, les travaux de Vannote ont très vite emporté une large approbation de la communauté scientifique et le concept de continuum fluvial est devenu le modèle de référence pour la description de la biocénose des écosystèmes lotiques. Il a notamment le mérite d’expliquer correctement certaines découvertes antérieures, comme celles de Robert Lauterborn (1918) et de Joachim Illies (1961).
Actuellement, on a surtout recours au modèle de Vannote pour diagnostiquer les perturbations des cours d’eau en comparant la composition réelle de sa biocénose à la composition théorique dans le modèle du continuum fluvial.
Études de la fragmentation écologique de cours d'eau
Les modèles d'estimation de perméabilité écologique d'un cours d'eau sont encore à affiner, et tout modèle du processus écologiques de dispersion doit toujours être associé à un plan d'échantillonnage adapté (modèle d'observation appuyé sur un échantillonnage cohérent[5], assez étendu (dans le temps et dans l'espace), avec une pression d'échantillonnage suffisante et n'interférant pas ou peu avec le comportement migratoire des organismes suivis).
En effet, il existe des facteurs climatiques et saisonniers, ainsi que des facteurs individuels et génétiques pouvant très significativement modifier les comportements de déplacements d'organismes tels que les poissons et crustacés[6].
Il existe aussi de nombreux facteurs discrets de fragmentation (dont thermiques, physico-chimiques, pollution...). De plus les causes de fragmentation d'un corridor aquatique sont encore mal cernées[5] ; certaines espèces pourraient ainsi être sensibles aux vibrations ou à certains champs électromagnétiques qui nous sont imperceptibles, ou encore à l'éclairage nocturne, tous éléments pouvant être fragmentant pour les espèces qui y sont sensibles (certains poissons notamment)...
Pour mieux étudier ou tenter modéliser les phénomènes de dispersion biologique et/ou de fragmentations naturelles et ou artificielles d'un continuum fluvial, on cherchera donc à :
- intégrer les dynamiques naturelles ou contextuelles d'extinction-colonisation-recolonisation propre à l'espèce ou au milieu[5], à plusieurs échelles au sein de la métapopulation[7] ;
- intégrer les impacts écosystémiques de l'éventuelle disparition d'autres espèces (prédateurs, ou espèce-fourrage, ou espèce symbiote ou espèce concurrente dans la niche écologique) et de l'éventuelle apparition d'autres espèces (espèce invasive ou parasite ou pathogène introduit par exemple) ;
- repérer et cartographier des voies et « noyaux de dispersion » active (loi (distribution) de Laplace modifiées) ou passive et/ou on mesure in situ, par comptage avec suivi par marquage-recapture les dispersions des individus du groupe étudié[5] ;
- intégrer des données anciennes (écologie rétrospective) ou sur l'écopotentialité du milieu, quand cela est possible. La position de l'espèce dans son aire de répartition est aussi à pendre en compte, de même qu'une éventuelle modification de cette aire en cours dans le contexte du dérèglement climatique.
Ainsi, à titre d'exemple, l'omble de fontaine (Salvelinus fontinalis) a été suivi de la sorte au Québec dans quatre cours d'eau traversés par une autoroute. Les recaptures ont dans ce cas montré une hétérogénéité marquée du comportement de dispersion au sein de la population : un groupe (48-72 % de tous les individus) était à dominante très sédentaire (caractérisée par des individus à distance moyenne de dispersion moyenne très courte ; de moins de 10 m)[8]. Un groupe secondaire était nettement plus mobile, caractérisé par une moyenne de dispersion de 56 à 1 086 m). L'étude n'a néanmoins pas pu mettre en évidence de preuve d'un véritable effet-barrière de l'autoroute sur la dispersion à travers les passages sous-routiers.
Limites du concept et propositions alternatives

Malgré ses succès, le concept de continuum fluvial, tel que développé aujourd'hui a un champ d’application limité :
- il a été développé pour certaines zones biogéographiques (tempérées à moyennement froides) ;
- il ne permet pas de décrire les portions stagnantes des cours d’eau (lacs, barrages, bras morts…) ;
- il ne tient pas ou peu compte des phénomènes de crue ;
- il présuppose que la végétation rivulaire et le contexte écologique du proche bassin-versant est « intact » ou peu modifié par l'homme, ce qui est rarement le cas ;
- il permet mal de prédire tous les impacts de la pollution (de l'air, des sols et des eaux);
- il ne tient pas ou peu compte de l'impact des affluents.
Pour tenter d’apporter des réponses à ces problèmes, le concept a été retravaillé par différents chercheurs.
- J.V. Ward et J.A. Stanford ont par exemple élaboré le Serial Discontinuity Concept (1983) pour tenir compte des ralentissements des eaux courantes puis l’Hyporheic Corridor Concept (1993) pour intégrer les dimensions latérales et verticales (rives et profondeur) à la problématique.
- Le Flood Pulse Concept de W.J. Junk et al. (1989), modifié par P.B. Bayley (1990) et K. Tockner et al. (2000) a encore amélioré la prise en compte des phénomènes d’inondation.
- Le cas des très grands fleuves est abordé par le Riverine Productivery Model de J.H. Thorp et M.D. Delong (1994).
- Le Telescoping Ecosystem Model de Fisher et al. (1998) permet d’expliquer les flux de matière entre les eaux de surface du cours d’eau proprement dit, les eaux de sub-surface (zones hyporhéique et parafluviale) et la zone riveraine.
Par ailleurs cette notion est complétée par la dynamique fluviale, qui est l'approche physique de l'équilibre et de l'évolution d'un cours d'eau.
Bibliographie
River Continuum Concept
- Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE (1980): The River Continuum Concept, Canadian Journal of Fisheries and Aquatic Sciences 37(1): 130-137. Texte intégral
- Wasson G. (1989): Eléments pour une typologie fonctionnelle des eaux courantes. Revue critique de quelques approches existantes, Bulletin d'écologie 20(2): 109-127.
- Maamri A., Chergui H., Pattee E. (1995): Impact des apports végétaux allochtones sur la distribution de la faune benthique dans un cours d'eau montagnard marocain, l'oued zegzel, Ecologie 26(2): 81-94.
Autres approches
- Junk JW, Bayley PB, Sparks RE (1989): The flood pulse concept in river floodplain systems. Canadian Special Publications of Fisheries and Aquatic Sciences 106: 110-127
- Ward JV, Stanford JA (1983): The serial discontinuity concept of river ecosystems. In: Fontaine TD, Bartell SM (Hrsg): Dynamics of lotic ecosystems, 29-42, Ann Arbor Science Publications, Ann Arbor, Michigan.
- Stanford JA, Ward JV (1993): An ecosystem perspective of alluvial rivers: connectivity and the hyporheic corridor, Journal of the North American Benthological Society 12, 48-60
- Bayley PB (1990): The flood pulse advantage and the restoration of river - floodplain-systems, Regulated Rivers: Research & Management 6, 75-86
- Tockner K, Malard F, Ward JV (2000): An extension of the flood pulse concept, Hydrological Processes 14, 2861-2883
- Thorp JH, Delong MD (1994): The riverine productivity model: A heuristic view of carbon sources and organic processing in large river systems, Oikos 70, 305-308
- Fisher, S. G., Grimm, N. B., Marti, E., Holmes, R. M. & Jones, J. B. (1998): Material spiraling in stream corridors : a telescoping ecosystem model, Ecosystems, 1 : 19-34.
- Bravard et Petit (2000) : "Les Cours d'Eau, Dynamique du Système Fluvial", PUF.
- Amorros et Petts (1994) : "Hydrosystèmes Fluviaux"
- Bravard, J. P. (1991). La dynamique fluviale à l'épreuve des changements environnementaux: Quels enseignements applicables à l'aménagement des rivières?. La Houille Blanche, (7-8), 515-522 (http://www.shf-lhb.org/articles/lhb/abs/1991/06/lhb1991047/lhb1991047.html résumé])
Notes et références
- Janine Gibert, Pierre Marmonier, Marie-José Dole-Olivier, Sous les eaux vives prospère un univers de curiosités biologiques ; Un fleuve peut en cacher un autre, La recherche, no 288, juin 96. - P. 44-46.
- Ovidio M., Philippart J. - C., Orban P., Denoël P., Gilliquet M., Lambot F. ; 2009; . Bases biologiques et éco-hydrauliques pour la restauration de la continuité piscicole en rivière : premier bilan et perspectives. Forêt Wallonne 101 : 18-29 (12 p., 7 fig., 1 tab., 13 réf.)
- F. Barthélemy, Les lamproies, Le Cordon Bleu, no 685, 15 décembre 1912
- Pringle, C. M. (1997). Exploring how disturbance is transmitted upstream: going against the flow. Journal of the north american Benthological society, 425-438. (résumé)
- McGeoch MA, Gaston KJ., Occupancy frequency distributions: patterns, artefacts and mechanisms ; Biol Rev Camb Philos Soc. 2002 Aug; 77(3):311-31 (résumé)
- Hawkes C., Linking movement behaviour, dispersal and population processes: is individual variation a key ? ; J Anim Ecol. 2009 Sep; 78(5):894-906. Epub 2009 Mar 6.
- Heatherly T, Whiles MR, Gibson DJ, Collins SL, Huryn AD, Jackson JK, Palmer MA, Stream insect occupancy-frequency patterns and metapopulation structure. Oecologia. 2007 Mar; 151(2):313-21. Epub 2006-11-08.
- Pépino M, Rodríguez MA, Magnan P (2012), Fish dispersal in fragmented landscapes: a modeling framework for quantifying the permeability of structural barriers ; Ecol Appl. 2012 Jul;22(5):1435-45. (résumé).
Voir aussi
Articles connexes
Liens externes
- (en) Wiki consacré à la restauration des rivières en Europe (522 exemples dans 31 pays), créé dans le cadre du européen RESTAURE.