Uranocentrodon
Uranocentrodon senekalensis

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Classe | Amphibia |
| Ordre | † Temnospondyli |
| Sous-ordre | † Stereospondyli |
| Famille | † Rhinesuchidae |
- Myriodon senekalensis van Hoepen, 1911 (préoccupée)
Uranocentrodon (littéralement « palais aux dents ») est un genre éteint d'amphibiens temnospondyles appartenant à la famille également éteinte des rhinésuchidés, ayant vécu durant la toute fin du Permien (Changhsingien), dans ce qui est aujourd'hui l'Afrique du Sud. Une seule espèce est connue, Uranocentrodon senekalensis, nommé à l'origine Myriodon par le paléontologue E.C.N. van Hoepen en 1911, avant d'être transféré par la suite dans un nouveau genre en raison de la préoccupation du nom en 1917.
Connu à partir d'un crâne mesurant 50 centimètres de long, Uranocentrodon était un grand prédateur ayant une longueur estimé jusqu'à 3,75 mètres[1]. Il a été proposé que ce taxon serait un probable synonyme de Rhinesuchus, mais cela n'est pas largement soutenu. Il était également considéré à l'origine comme ayant vécu durant le Trias, mais une analyse plus récente place son âge juste en dessous de la limite Permien-Trias[2].
Découverte et spécimens
Les fossiles de l'animal maintenant connu sous le nom d'Uranocentrodon sont découverts pour la première fois dans une carrière de grès de la formation de Normandien (en) près de la ville de Senekal, dans la province de l'État libre, en Afrique du Sud. Le premier exemple de ces fossiles est un crâne fragmentaire excavé par une famille locale. Des fouilles et des recherches ultérieures entreprises par diverses parties mettent à jour d'autres spécimens provenant de la carrière. Parmi ces spécimens figurent trois squelettes presque complets se chevauchant. Le crâne initial est déterminé comme appartenant au même individu que l'un des squelettes. Le crâne et les trois squelettes sont acquis par le musée du Transvaal et reçoivent les désignations TM 75, TM 75d et TM75h par le paléontologue E.C.N. van Hoepen. En 1911, il donna le nom officiel Myriodon senekalensis aux spécimens au moyen d'une brève description publiée en néerlandais. Il publie une description plus complète quelques années plus tard, en 1915[1]. Deux crânes supplémentaires sont mentionnés dans cette description, bien que leur localisation soit actuellement inconnue. En 1917, il est déterminé que le nom Myriodon est déjà utilisé par un genre de poisson et qu'un nouveau nom devait être utilisé. Ainsi, van Hoepen renomme le taxon Myriodon senekalensis en Uranocentrodon senekalensis, ce qui se traduit littéralement par « le palais aux dents piquantes du Senekal »[3]. Les spécimens originaux du musée du Transvaal sont collectivement appelés les syntypes (en) d'Uranocentrodon[2].
Au fil du temps, des nouveaux fossiles supplémentaires sont découverts. À peu près au même moment où les spécimens du Transvaal sont décrits, Robert Broom découvrit des fragments de crâne qu'il considérait comme appartenant à une nouvelle espèce de Rhinesuchus, Rhinesuchus major[4]. Cependant, les chercheurs pensent maintenant que ce crâne, désigné 60C 1-1a, appartient à Uranocentrodon, faisant de R. major un synonyme junior du genre[2].
En 1915, Sidney H. Haughton, décide de synonymiser Uranocentrodon (à l'époque encore appelé Myriodon) avec Rhinesuchus, bien qu'il le garde comme une espèce distincte au sein du genre. Il décrit également un autre squelette pratiquement complet, y compris du matériel crânien, conservé au musée national de Bloemfontein, maintenant catalogué NMQR 1483. Il mentionne également un fragment de mâchoire, maintenant catalogué SAM-PK-2783[5]. La plupart des autres analyses considèrent qu'Uranocentrodon est distinct de Rhinesuchus[2]. D'autres restes connus incluent TM 185 (un crâne décrit par Broom en 1930), TM 208 (un membre antérieur droit) et CGP 4095 (un autre crâne)[6].
Paléobiologie
Branchies
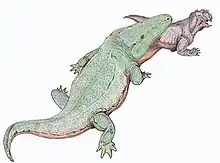
La préservation des squelettes originaux du Transvaal permet de préserver des parties délicates de ces derniers. Parmi ces parties, on compte environ trois rangées d'os minuscules (osselets branchiaux) recouverts de fines structures semblables à des dents (denticules branchiaux). Ces structures apparaissent près du cou de l'un des squelettes et presque certainement attachées aux arcs branchiaux des branchies pendant que l'animal était vivant[1]. Bien que de tels os soient rares parmi les stéréospondyles et inconnus chez tous les autres rhinesuchidés, cela peut simplement être dû au fait que les os d'autres genres sont préservés dans des sédiments plus rugueux où ces os délicats pourraient être fragmentés ou difficiles à trouver[7].
Bien qu'il soit évident qu'Uranocentrodon aurait eu des branchies, il est difficile de déterminer de quel type de branchies il s'agit. D'une part, il pourrait s'agir de branchies internes (en) comme celles des poissons, peu visibles de l'extérieur du corps. D'autre part, ils auraient pu être des branchies externes en forme de tige comme celles des larves de salamandres modernes ou même des salamandres adultes néoténiques tels que le genre Necturus ou l'axolotl. Les branchies externes ont dû évoluer à partir des branchies internes au cours de l'évolution des amphibiens, bien que l'emplacement précis de cette transition soit controversé. Les os de soutien des branchies conservés chez les anciens amphibiens présentent de nombreuses similitudes avec ceux des branchies de poisson et des branchies des salamandres. Selon le groupe auquel les paléontologues comparent les os, différents types de branchies peuvent être déduits. Cette énigme, connue sous le nom de paradoxe de Bystrow, rendait difficile l'évaluation des branchies chez les anciens amphibiens tels que Uranocentrodon[8].
Le paradoxe de Bystrow sera finalement résolu par une étude datant de 2010. Cette étude révèle que les structures cérato-branchiales rainurées (composants des arcs branchiaux) sont corrélées avec les branchies internes. Les anciens tétrapodes qui conservent des cératobranchiaux rainurés, comme Dvinosaurus, n'ont probablement que des branchies internes à l'âge adulte. Néanmoins, les branchies externes sont directement préservées en tant que tissus mous chez certains temnospondyles. Cependant, ces situations ne se produisent que chez les spécimens larvaires ou les membres de groupes spécialisés tels que les branchiosauridés (en). Une espèce vivante de dipneuste (Lepidosiren paradoxa) a des branchies externes sous forme de larves qui se transforment en branchies internes à l'âge adulte. Bien que les spécimens de dvinosaures adultes aient des caractéristiques squelettiques corrélées aux branchies internes, certains spécimens larvaires d'un autre dvinosaure, Isodectes, conservent des tissus mous de branchies externes. Ainsi, le développement des branchies des dvinosaures (et vraisemblablement d'autres temnospondyles, comme Uranocentrodon) refléterait celui de Lepidosiren paradoxa. Bien que cette caractéristique soit probablement un exemple d'évolution convergente (car d'autres dipneustes possèdent exclusivement des branchies internes), elle reste un indicateur utile de la façon dont les branchies de temnospondyles se développaient. En conclusion, les branchies d'Uranocentrodon seraient probablement internes comme celles d'un poisson à l'âge adulte, mais externes comme celles d'une salamandre à l'état larvaire[8].
Armures dermiques
Certains des spécimens du Transvaal conserve également de grandes plaques d'écailles ou d'écailles osseuses autour du corps. Les écailles sur le ventre de l'animal sont disposées en rangées diagonales parallèles qui convergent vers la ligne médiane du corps et divergent lorsque les rangées s'étirent vers la queue. Chaque écaille a une crête au milieu, et les écailles plus loin vers la ligne médiane chevauchent celles qui sont plus éloignées. Le long de la ligne médiane, une rangée d'écailles plates et larges s'étendent de la gorge à la queue. Alors que ces écailles abdominales sont faites d'os, les écailles d'autres parties du corps ont moins de structure osseuse et seraient probablement faites de kératine à la place[1].
Les écailles sur les côtés du corps sont plus plates et plus petites que les écailles osseuses du ventre. Les écailles à l'arrière du corps étaient similaires, bien que de forme plus arrondie, avec quelques écailles plus grandes près de la ligne médiane. Les écailles des membres postérieurs et le dessous de la région des hanches sont similaires à celles du dos, bien qu'aucun tégument ne soit conservé sur les membres antérieurs ou la queue. Ainsi, il est probable qu'au moins la queue serait sans armure et uniquement recouverte de peau nue[1].
Notes et références
Notes
Références
- (en) E.C.N. Van Hoepen, « Stegocephalia of Senekal, O.F.S », Annals of the Transvaal Museum, vol. 5, no 2, , p. 125-149 (DOI 10.10520/AJA00411752_812
 , S2CID 134584405, lire en ligne [PDF])
, S2CID 134584405, lire en ligne [PDF]) - (en) C.A. Marsicano, E. Latimer, B. Rubidge et R.M.H. Smith, « The Rhinesuchidae and early history of the Stereospondyli (Amphibia: Temnospondyli) at the end of the Palaeozoic », Zoological Journal of the Linnean Society, vol. 181, no 2, , p. 357–384 (DOI 10.1093/zoolinnean/zlw032 , S2CID 89770655)
- (en) E.C.N. Van Hoepen, « Note on Myriodon and Platycranium », Annals of the Transvaal Museum, vol. 5, no 4, , p. 217 (DOI 10.10520/AJA00411752_837 , S2CID 125639445, lire en ligne [PDF])
- (en) R. Broom, « Note on the temnospondylous stegocephalian, Rhinesuchus », South African Journal of Geology, vol. 14, no 1, , p. 79–81 (DOI 10.10520/AJA10120750_1429 , S2CID 135050076, lire en ligne [PDF])
- (en) S.H. Haughton, « On the genus Rhinesuchus, with notes on the described species », Annals of the South African Museum, vol. 22, , p. 65–77 (lire en ligne)
- (en) R. Broom, « Notes on some Labyrinthodonts in the Transvaal Museum », Annals of the Transvaal Museum, vol. 14, no 1, , p. 1–10 (DOI 10.10520/AJA00411752_643 , S2CID 195022656, lire en ligne [PDF])
- (en) Rainer R. Schoch, « The evolution of metamorphosis in temnospondyls », Lethaia, vol. 35, no 4, , p. 309–327 (ISSN 1502-3931, DOI 10.1111/j.1502-3931.2002.tb00091.x, S2CID 83496782)
- (en) Rainer R. Schoch et Florian Witzmann, « Bystrow's Paradox - gills, fossils, and the fish-to-tetrapod transition », Acta Zoologica, vol. 92, no 3, , p. 251–265 (ISSN 1463-6395, DOI 10.1111/j.1463-6395.2010.00456.x, S2CID 85651134, lire en ligne)
Voir aussi
Articles connexes
Liens externes
- Ressources relatives au vivant :