Telema tenella
Telema tenella est une espèce d'araignées aranéomorphes de la famille des Telemidae[1].

| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Sous-embr. | Chelicerata |
| Classe | Arachnida |
| Ordre | Araneae |
| Sous-ordre | Araneomorphae |
| Famille | Telemidae |
| Genre | Telema |
Distribution
Cette espèce est endémique des Pyrénées. Elle se rencontre dans des grottes du massif du Canigou dans les Pyrénées-Orientales en France et de la province de Gérone en Catalogne en Espagne[1] - [2]. En France on la rencontre notamment vers Montferrer et La Preste dans diverses grottes et une mine.
Description
Telema tenella mesure de 1,5 mm à 2,5 mm. Cette espèce troglobie ne possède ni yeux ni poumons.
Son prosoma ou céphalothorax, fauve-rougeâtre clair, est dépourvu d'yeux et de strie thoracique médiane contrairement à celui des Leptonètes ; l’anophtalmie y rend imprécises les limites du bandeau ou clypeus. Il porte des chélicères divergentes et de longues pattes grêles, garnies de poils sensoriels et de glandes prétendues "d’Emerit » qui ne sont pas caractéristiques car présentes aussi chez d’autres Araignées. Le mâle possède un bulbe copulateur simple, arrondi, avec un style très court, en apophyse unguiforme (fig.1) et les femelles, un réceptacle séminal impair, médian, remarquablement volumineux qu'ont aussi décrit Chunxia Wang, Ribera Carles et Shuqiang Li (2012).

L’abdomen est court, globuleux, avec une région épigastrique convexe, deux paires de stigmates trachéens ventraux, un colulus, six filières terminales et présente surtout une étrange coloration d’un bleu-vert "bouteille" soulignée par divers auteurs (Simon, 1882 ; Fage, 1913 ; Lopez,1976[3],1977[4],1980[5]), retrouvée d’ailleurs chez Usofila pecki (Nouvelle Calédonie : Brignoli, 1980), mais avec une nuance « émeraude" . Dans ses premières coupes histologiques de Telema tenella, A.Lopez (1976)[3] a constaté que cette coloration particulière dépend de l’intestin moyen, plus précisément de ses diverticules chylentériques à large lumière polycyclique contenant des "inclusions" (sphérites) de même teinte.
Par ailleurs, l'abdomen renferme de grandes glandes coxales (Lopez & al.,1983)[6], un appareil séricigène original (Lopez,1983)[7], chez le mâle des canaux déférents très développés qui élaborent les spermatophores (Lopez & al.,1981) [8]et, chez la femelle, un réceptacle séminal impair, médian remarquablement volumineux, vu aussi par Chunxia Wang, Ribera Carles et Shuqiang Li (2012) et qui présente une structure microscopique très particulière (Lopez,1983)[9].
Intérêt
Telema tenella présente non seulement un intérêt patrimonial en tant que troglobie, endémique et "fossile vivant" mais aussi un intérêt anatomo-physiologique majeur par la production et l'usage de spermatophores ( A. Lopez,1976[3],1977[4]), une étude histologique et ultrastructurale complémentaire (Lopez & Juberthie, 1980)[5] ayant également confirmé la présence de sphérocristaux ou sphérites intestinaux responsables de la coloration abdominale. L'existence de spermatophores (Fig.2), dont l'ultrastructure est aujourd'hui bien connue chez Telema (Lopez & al., 1981)[8], a été retrouvée dans le cas d'autres Telemidae (Lopez & Legendre, 1978)[10].Chunxia Wang, Ribera Carles et Shuqiang Li, 2012, n'en font aucune mention. Ils ne signalent pas de spermatophores ni dans le bulbe du mâle où ils sont pourtant présents après induction (Fig.2), ni dans la spermathèque de la femelle où ils sont également visibles après copulation.
Cet article souligne toute l'importance de l'anatomie microscopique dans la recherche de nouveaux organes, une meilleure compréhension de certains traits du comportement animal et l'apport de critères plus rigoureux dans la systématique.
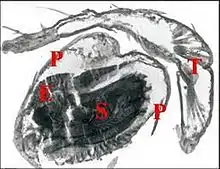
Comportement

Telema tenella installe sa toile dans des « niches » très variées du milieu hypogée : anfractuosités des piliers, bornes et coulées stalagmitiques, interstices de murets et même dans les trous de barre à mine ! Cette toile est une nappe ténue (Fig.3), très légère, d’aspect non visqueux, à petites mailles irrégulières, haubanée par quelques fils tenseurs, subhorizontale, d’étendue variable, pouvant dépasser 15 cm de long, et à concavité inférieure plus ou moins marquée, où l’Araignée se tient ventre en l’air.
La femelle édifie des cocons ovigères qui, d'après les observations sur le terrain de Lopez et celles de Juberthie (1985) dans ses élevages, ont une forme discoïdale en lentille plan convexe (Fig.4). Ils sont très petits, mesurent de 1,5 à 1,8 mm de diamètre, sont fixés sur les concrétions adjacentes ou dans un lacis de fils périphériques tendant la toile et y adoptent une disposition aléatoire, de verticale à horizontale. La coexistence habituelle de plusieurs d'entre eux serait en rapport avec l'espacement des pontes dans le temps et la lenteur du développement embryonnaire. Chaque cocon ne contient que très rarement un seul œuf, particularité admise comme constante à la suite de Fage (1913 : pl.69, fig.21), en fait beaucoup plus souvent deux, et surtout trois ou quatre (Juberthie,1985). Ces œufs ont un diamètre très réduit, d'environ 0,45 mm. Une même femelle construit chaque année un nombre moyen de quatre cocons, un seul à chaque ponte.
En ce qui concerne le cycle vital, Juberthie (1985) a pu constater, toujours dans ses élevages au Laboratoire souterrain de Moulis, que la durée du développement embryonnaire, depuis la ponte jusqu'à la mue suivant l'éclosion, est de l'ordre de 9 à 11 mois, donc exceptionnellement lente et, sans aucun doute, la plus longue connue à ce jour chez les Invertébrés souterrains troglobies. Le développement post-embryonnaire, depuis la sortie du cocon jusqu'à la mue imaginale, est de l'ordre de 3 ans. Quant à la durée de vie adulte, du moins chez la femelle, elle peut être estimée à une douzaine d'années. Il s'ensuit que Telema tenella a une espérance totale de vie d'environ 16 ans, longévité extraordinaire pour un si petit animal !

Anatomie interne
Les premières coupes histologiques de Telema tenella effectuées par André Lopez[3] ont confirmé l’absence complète d’yeux, avec une régression cérébrale corrélative des centres visuels. Elles ont montré aussi deux glandes à venin banales, des glandes glandes gnathocoxales ou " salivaires " sans dimorphisme sexuel, de gros néphrocytes, du tissu réticulé, un réseau trachéolaire dense, de grosses glandes coxales et surtout, un appareil génital très particulier ainsi que des sphérites intestinaux lui donnant sa couleur abdominale particulière. Enfin, les glandes séricigènes présentent aussi des caractères originaux.
Glandes coxales
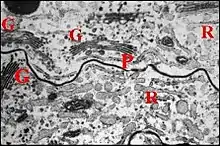
Au nombre d'une paire, elles s’ouvrent aux hanches des P I. Leur grand labyrinthe s’étend le long du cerveau jusqu’à la partie postérieure du prosoma (Fig.).
Ce labyrinthe a une "striation" typique dont l'étude a été effectuée ultérieurement au microscope électronique à transmission (M.E.T). ([null Lopez,1983d]). Elle est liée à des replis membranaires qui découpent profondément le hyaloplasme basal, très riche en mitochondries, et sont solidarisés par de belles jonctions septées ("desmosomes cloisonnés") (Fig.)
Appareil génital
C'est le tractus génital, femelle et surtout mâle,qui représente la particularité la plus singulière de cette Araignée pyrénéenne[3] - [4], à un degré moindre du genre Apneumonella (Afrique)[10] et, probablement aussi, des autres Telemidae.
Appareil génital mâle
Il comporte une paire de testicules subsphériques et deux canaux déférents longs, très sinueux, convolutés, s’unissant sur la ligne médiane en un conduit terminal commun ouvert au gonopore, dans le sillon épigastrique.
Canaux déférents
L’ensemble des canaux est comparativement plus gros et surtout plus épais que dans les autres familles d’Araignées,. Avec son contenu, il forme un massif volumineux, long d’environ 250 µm chez un mâle de 1,3 mm et occupe ainsi toute la partie antéro-ventrale de l’abdomen . Chaque déférent contient une substance amorphe, des spermatozoïdes et, dans sa partie distale ou terminale différenciée, une formation tubuleuse brun-jaunâtre, large d’environ 18 µm, triangulaire en coupe transversale, évoquant un «étui» et pourvue de grêles expansions latérales. Les gamètes se logent dans ce spermatophore, superposés en « pile d’assiettes » ou groupés comme les érythrocytes dans un capillaire de Vertébré. Revêtant avec ses expansions un aspect caricatural de «Myriapode », le spermatophore se retrouve, apparemment tel quel, dans les coupes histologiques du palpe mâle, , plus précisément de son bulbe et, chez la femelle, dans la spermathèque. Il diffère de celui d’Apneumonella sp., autre araignée Telemidae, qui est jaune-rougeâtre, lové sur lui-même, contient aussi des gamètes empilés « comme les comprimés dans leur étui »mais a un aspect cylindrique, une cavité axiale régulière et ne présente aucune expansion (Lopez,1978[10]).
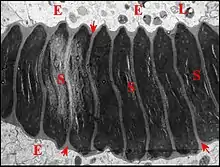
Histologie
La structure du déférent n’est pas uniforme. Ce conduit montre en effet une portion proximale sinueuse dont l’épithélium, très bas, rappelle un revêtement endothélial, une portion intermédiaire à cellules plus hautes et une partie distale longue et complexe. Renfermant le spermatophore, cette dernière est formée par des cellules prismatiques beaucoup plus grandes, surtout ventralement (hauteur : 30 µm) et à noyaux ovoïdes très apparents.
Ultrastructure
Outre son épithélium interne, la paroi du déférent montre au microscope électronique à transmission (M.E.T.) une lame basale et une couche de fibres musculaires dont le hyaloplasme contient des faisceaux de myofibrilles caractéristiques isolés.L'épithélium est simple, prismatique et sécrétoire. Ses cellules sont solidarisées par des zonulae adherens sub-apicales et de longues jonctions septées. Sa lumière est large, arrondie et régulière dans la portion proximale, puis ovale au début de la partie différenciée ou portion intermédiaire (Fig.), pourvue enfin d’une gouttière ventrale lui
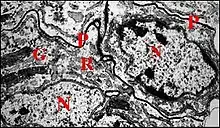
conférant un curieux aspect en « as de pique » dans le reste de la partie différenciée ou portion distale. Il s’ensuit qu’au niveau de cette dernière la paroi déférentielle présente une zone dorsale mince et une zone ventrale ou accessoire plus épaisse (Schéma-diagramme).
Dans la portion proximale, en continuité avec le testicule, les cellules épithéliales aplaties ont un plasmalemme basal présentant de nombreux replis. Leurs sommets sont réunis par des jonctions sub-apicales. Le cytoplasme renferme des lysosomes (Fig.). Le réticulum endoplasmique est granulaire, très abondant et produit de petites vésicules s'ouvrant dans la lumière pour y déverser leur contenu sécrétoire. Ce même matériel englobe les spermatozoïdes encore dépourvus de coque ou capsule d'enkystement, et, lorsque ces derniers s’empilent, s’insinue entre leurs faces juxtaposées pour y former des disques plus opaques apparaissant comme des bandes régulières (600 A) dans les coupes longitudinales (Fig.).
Dans la partie différenciée, les cellules épithéliales sont plus hautes, uniformément (portion intermédiaire), et surtout dans
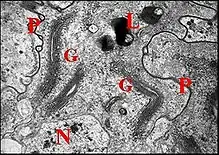
la zone ventrale de la grande portion distale (Schéma-diagramme). Leur noyau central renferme une chromatine fine et dispersée (Fig.). Le réticulum endoplasmique est formé par de nombreuses cisternae lisses et granulaires (Fig.16,18) dont dérivent des vésicules à contenu peu contrasté. L'appareil de Golgi est remarquablement développé (Fig.). Il se compose d’empilements sacculaires ou dictyosomes surtout nombreux dans la zone dorsale et élaborant une grande quantité de vésicules à contenu opaque.Les mitochondries sont peu nombreuses et pourvues de crêtes parallèles. Les vésicules golgiennes et réticulaires libèrent leur contenu dans la lumière au niveau des pôles apicaux. Il s’y fusionne avec la sécrétion provenant de la partie proximale pour former un matériel granuleux positif à l’APS et à la tétrazoréaction de Danielli, donc glycoprotéique[8].
Les spermatozoïdes mûrs se sont formés dans le testicule à partir de spermatides d’abord polymorphes et amiboïdes, puis polarisées, pourvues d’un noyau allongé, enfin flagellées[8] . Chacun d’eux est entouré par une capsule ou coque
d'enkystement glycoprotéique (Fig.) semblable à celle des autres Araignées et peu condensé. Son cytoplasme est abondant, aucune phase d’élimination cytoplasmique n’ayant été clairement observée au cours de la spermiogénèse ; il est
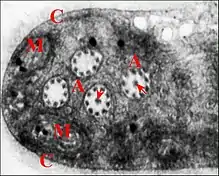
très riche en mitochondries à crêtes concentriques (Fig.), en vésicules ovoïdes et en membranes enchevêtrées d'origine golgienne vraisemblable. Le noyau et la baguette acrosomienne sont étirés et enroulés sur eux-mêmes, le premier suivant 1 tour et demi ou 2. L'axonème du flagelle, de type 9 + 3 comme chez les autres Araneides, est rétracté dans le cytoplasme où il s’enroule sur 4 à 5 tours (Fig. ). De plus, chaque gamète montre un « prolongement » qui pénètre dans une digitation, y est accompagné par la capsule et renferme lui aussi des organites (vésicules, , amas parallèles de membranes sinueuses, (Fig.).
Description
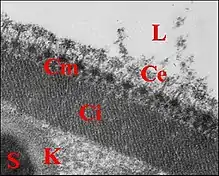
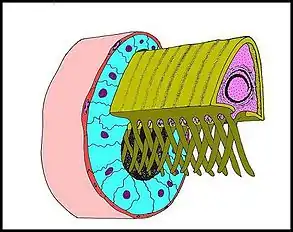
Le spermatophore s'individualise dans la portion distale du canal déférent. ... Il a bien la forme insolite d’un étui prismatique triangulaire (Schéma), avec deux grandes faces latérales un peu convexes et une face basale plus réduite, légèrement concave. Il est ouvert à ses deux extrémités et selon une génératrice, au niveau de la même base qui montre donc une déhiscence longitudinale lui conférant aussi un aspect en gouttière (Schéma-diagramme , Fig.). Les rebords de cette fente portent chacun une série de digitations disposées en alternance, initialement sur un rang puis sur 2 files parallèles, décalées mais s’entrecroisant ( Schéma ). Ces digitations contiennent les prolongements des spermatozoïde La paroi du spermatophore est elle-même formée par 3 couches superposées, interne, moyenne et externe à l’exception toutefois des bords basaux (Fig.) et des digitations (Fig.) qui n’en ont qu’une seule, l’interne [9]Lopez,1981c).Cette couche interne est la plus épaisse (0,45 µm) et possède une structure paracristalline (Fig.). Elle semble striée dans les coupes transversales (Fig.) mais montre dans les longitudinales et axiales un réseau polygonal très régulier,

en «nid d’abeilles», dont chaque élément constitutif est centré par une fibre dense. La distance moyenne entre 2 fibres est de 75 Å (Fig.). La couche moyenne ou intermédiaire est beaucoup plus mince (500 à 600 Å) et forméepar des agrégats de matériel dense lui donnant un aspect hétérogène alvéolé (Fig.). La couche externe, qui est la plus originale, se présente comme un ensemble de piliers étroits (diamètre = 150 Å), d’abord bas et granuleux dans la paroi en formation (Fig.) puis plus hauts (0,2 µm) et homogènes (Fig.). Ils s’alignent en séries parallèles perpendiculairement à l’axe de la gouttière et sont ainsi responsables d’une striation transversale.
Cette ultrastructure est retrouvée dans la spermathèque lorsque le spermatophore y a été introduit (Fig.), d'abord inchangée (Fig.) puis avec des modifications portant sur la couche interne (aspect "flou") et la couche externe (inclinaison des piliers un même sens) (fig.). La couche moyenne parait plus nette tandis que la capsule ou coque d'enkystement gamétique semble s'effacer(Fig.).
Le "prolongement" de chaque spermatozoïde pénètre avec sa capsule dans une digitation formée par la seule couche
interne et qui l'englobe comme un petit étui individuel (Fig.).
Commentaires
Sur le plan anatomique, le canal déférent de Telema tenella est remarquable par son volume, son intense activité sécrétoire d’origine golgienne et le fait qu’il élabore une structure ayant bien tous les caractères d’un spermatophore. Bien que ce dernier ne soit pas un produit de glandes annexées à l’appareil génital et n’ait pas la forme de capsules closes, plus ou moins complexes, souvent pédiculisées comme chez les autres Arthropodes (dont les Scorpiones et Pseudoscorpions parmi les Arachnides), il n’en présente pas moins les particularités essentielles de ce type d’ « appareil ». Il s’agit en effet d'une formation autonome, bien individualisée, régulière et géométrique, en gouttière digitée, possédant une paroi propre à ultrastructure complexe, monolaminaire dans sa face basale et les digitations, trilaminaire ailleurs, avec une couche paracristalline et les curieux "piliers" externes. « piliers » externes. Une telle organisation fait défaut chez toutes les autres

Araignées, y compris celles qui présentent des « aggrégats » de spermatozoïdes comme les Dysderidae ("sphérulation" :Lopez,1972) (Fig. ), les Filistatidae et même certains Mygalomorphes dont les gamètes se groupent à plusieurs dans une même coque d'enkystement (Nemesia, M.E.T.:Lopez,1981). Décrivant de semblables "aggrégats" dans le cadre de ses "coenospermies", Alberti (1988) y a inclus aussi les spermatophores des Telemidae, considérés par lui comme une simple forme « complexe » des précédentes. Les ultrastructures observées chez Telema tenella invalident définitivement l'opinion critique de ce dernier auteur ainsi que ses assertions ultérieures, en collaboration, sur la même espèce et sa famille (Albert & Coyle,1991; Michalik & Alberti,2004), toutes ses images différant des photos ci-contre . Les spermatozoïdes empilés dans le spermatophore ont des caractères semblables à ceux des autres Araignées (capsule glycoproteinique individuelle, triplet axial, rétraction de l’axonème enroulé comme le noyau), mais en diffèrent néanmoins parce qu’ils conservent tout le cytoplasme de la spermatide.
Sur le plan fonctionnel,le spermatophore représente pour les gamètes mâles un «conditionnement» ou appareil d’ «emballage» renforçant leur protection déjà assurée par les capsules individuelles tout en leur permettant de progresser vers l’aval et le gonopore. Ce cheminement dans le déférent pourrait être facilité non seulement par la contraction des muscles pariétaux, par l'"accrochage" des digitations à la paroi du canal bien qu’elles ne présentent pas d’ultrastructure motrice et par la striation transversale jouant également un rôle mécanique. Le spermatophore est initialement produit par le canal déférent sur un mode ininterrompu, semble bien ouvert à ses deux extrémités et doit donc subir un « tronçonnement » ultérieur lors de la « transmission spermatique ». Bien qu’il n’ait jamais été observé, ce processus existe sans aucun doute . Lors de l’« éjaculation primaire », des fragments issus du gonopore pourraient être déposés sur une toile spermatique dépourvue de [null sécrétion épigastrique] et absorbés ensuite dans les bulbes (induction spermatique). L’apophyse unguiforme de ces derniers, leur concavité et l’orifice en fente extensible permettraient le recueil puis la pénétration des fragments dans les tubes séminifères où ils se pelotonnent jusqu’à l’accouplement (Fig . 5, 6)et leur insertion ultérieure (« éjaculation secondaire ») dans la [null spermathèque femelle].Coyle,1991
Sur le plan phylogénique, l’élaboration et l’usage d’un spermatophore chez Telema tenella et probablement aussi chez les autres Telemidae apparaissent comme le maintien d’un caractère archaïque. Alexander et Ever (1957) avaient d’ailleurs envisagé son existence chez les Araignées primitives et sa perte ultérieure lors de l’évolution dans le tissage de la toile. Cette dernière hypothèse ne paraît toutefois pas applicable à Telema car elle construit des cocons ovigères et un édifice soyeux en nappe assez perfectionnés. Par ailleurs, et comme déjà évoqué par Lopez (1977) bien qu'il ne connaisse pas encore Telema tenella , le spermatophore des autres Araignées pourrait avoir été remplacé, en cours d’évolution, par le produit sécrétoire des glandes prégonoporales (appareil épigastrique) qui serait alors son équivalent.
Sur le plan systématique, il apparait que grâce au spermatophore, les genres Telema et Apneumonella peuvent être rapprochés l’un de l’autre dans le cadre d’une parenté étroite mise en doute par Lehtinen (1967) trop axé sur l’appareil visuel. La famille des Télémides se trouve ainsi validée, non seulement par la structure particulière des organes génitaux (Brignoli,1973) mais surtout par leur contenu. Elle est donc bien distincte de celle des Leptonetidae, auxquels Fage (1913) les rattacha naguère, car ces derniers possèdent un bulbe copulateur plus complexe, deux spermathèques symétriques et surtout, des gamètes mâles logés sans ordre apparent dans une substance amorphe inorganisée.
Sphérocristaux (Sphérites)
Les cellules à ferment et absorbantes de l'intestin renferment une foule d’ « inclusions » arrondies, de taille assez variable, colorées naturellement en bleu-vert (Fig. 5), paraissant « encapsulées » et opaques ou de structure réticulée , retrouvées dans la poche cloacale et seules responsables, par leur pullulation, de la teinte
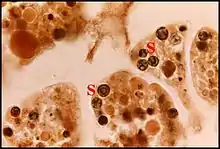
générale de l’abdomen, d'abord interprétées comme des « microorganismes …peut être des symbiontes particuliers, contenant un pigment à métal chromogène, nécessaires pour la survie précaire de Telema tenella » ( Lopez,1976)[3]. En fait, l’étude au microscope électronique à transmission montra plus tard ( Lopez,1980)[5] qu’il s’agit de sphérites ou sphérocristaux, concrétions inertes, polymorphes, rondes ou ovoïdes, zonées concentriquement, associant un « nucleus » opaque central et une série de strates périphériques alternativement claires et sombres (Fig.6). Ces sphérites naissent dans des vésicules du réticulum par condensation d’un matériel granuleux abondant : le « nucleus » y apparait en premier et s’entoure ensuite des strates concentriques dont le nombre semble augmenter avec l’âge et le volume de la concrétion. Leur analyse chimique sur coupes par microsonde à rayons X (W.Humbert, Strasbourg) y a mis en évidence du calcium (le plus abondant : 100 chocs par seconde), de l’aluminium (un élément pourtant rare dans ce type de concrétion : 10 chocs par seconde), du soufre, du phosphore, du potassium, mais pas de cuivre, pourtant présent dans le biotope et qui aurait pu être responsable de la

couleur verte des sphérites. Connus aussi chez les Nématodes, les Opilions et surtout, de très nombreux Insectes, dont les Collemboles, les sphérocristaux jouent un rôle dans les régulations ionique, hydrique et dans l'excrétion par accumulation. Il en est probablement de même pour ceux de Telema tenella.
Appareil séricigène
L'appareil séricigène offre lui aussi une activité sécrétoire originale (Lopez,1983 c).
Taxinomie
Cette espèce a été décrite en 1882 par l'arachnologiste français Eugène Simon (1848-1924). Elle est l'espèce-type du genre Telema.
Annexes
Bibliographie
- Eugène Simon, « Études Arachnologiques. 13e Mémoire. XX. Descriptions d'espèces et de genres nouveaux de la famille des Dysderidae », Annales de la Société entomologique de France, 6e série, vol. 2, , p. 201-240 (lire en ligne)
- (en) Chunxia Wang, Ribera Carles et Shuqiang Li, « On the identity of the type species of the genus Telema (Araneae, Telemidae) », ZooKeys, no 251, , p. 11–19 (DOI 10.3897/zookeys.251.3616).
- C. Juberthie et A. Lopez, « À propos du spermatophore et des sphérocristaux de l'araignée Telema tenella(Telemidae) : quelques données ultrastructurales », dans C. R. Vè. Colloque Arach. IX, 1979, Barcelone, (lire en ligne), p. 111-118
- A. Lopez et H Salvayre, « L'Araignée cavernicole pyrénéenne Telema tenella Simon et son habitat », Bulletin de la Société d'étude des Sciences naturelles de Béziers, vol. 4, no 45, , p. 17-23
Liens externes
- (en) Référence Animal Diversity Web : Telema tenella (consulté le )
- (en) Référence Fauna Europaea : Telema tenella Simon, 1882 (consulté le )
- (fr+en) Référence ITIS : Telema tenella Simon, 1882 (consulté le )
- (en) Référence World Spider Catalog : Telema tenella Simon, 1882 dans la famille Telemidae +base de données (consulté le )
Notes et références
- WSC, consulté lors d'une mise à jour du lien externe
- Entoflorachne, description de l'espèce Telema tenella par André Lopez-Moncet
- Lopez et Salvayre 1976.
- A. Lopez, « Sur un nouveau mode de reproduction chez les Araignées : existence de spermatophores chez Telema tenella Simon. Bull. Soc. Zool. France, 102, p. 261- 266. », Bull. Soc. Zool. France, 102, p. 261- 266.,
- Juberthie et Lopez 1980.
- A.Lopez, avec L.Juberthie-Jupeau et J.C.Bonaric., « Structure et ultrastructure des glandes coxales chez Telema tenella Simon (Araneae, Telemidae). », Mém.Biospéol.,X, p.433-437,
- A.Lopez avec J.Kovoor -, « Structure et ultrastructure de l'appareil séricigène chez Telema tenella. Simon (Araneae, Telemidae). Mém.Biospéol.,X, p.419-425. », Mém.Biospéol.,X, p.419-425.,
- A. Lopez, avec C. Juberthie et J. Kovoor, « Spermiogenesis and spermatophore in Telema tenella Simon (Araneae : Telemidae). An ultrastructural study. », Int. J.Invert. Reprod., 3, 181-191.,
- A.Lopez, avec L.Juberthie-Jupeau, « Structure et ultrastructure de la spermathèque chez Telema tenella. Simon (Araneae, Telemidae). », Mém.Biospéol.,X, p.413-418.,
- A. Lopez, avec R.Legendre, « Présence d’un spermatophore dans le genre Apneumonella (Araneae : Telemidae) : valeur taxonomique et problèmes de biologie sexuelle. », Bull.Soc.Zool.France, 103, p.35-41.,