Desmosome
Un desmosome est une région où la membrane plasmique d'une cellule adhère à une cellule adjacente ou à la lame basale sous-jacente, il est alors appelé hémidesmosome. La nature des filaments intermédiaires est variable (cytokératine dans les cellules épithéliales, vimentine dans les fibroblastes et les endodermes, neurofilament dans les cellules nerveuses ou encore lamine dans les noyaux).
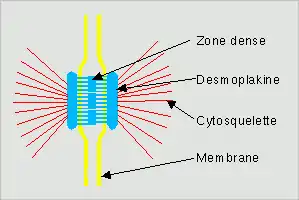
Structure du type macula adherens
Les desmosomes du type macula adherens, se retrouvant presque exclusivement dans les cellules épithéliales par les filaments intermédiaires associés, sur leur extrémité COOH intracytoplasmique, à des cadhérines. Ainsi les microfilaments intermédiaires ne sont pas liés les uns aux autres, ils sont ancrés dans une plaque cytoplasmique dense, constituées de desmoglobine (lié au filaments) et de desmoplakine (liés à la desmocolline ou la desmogléine) grâce à ces cadhérines. l'espace intercellulaires entre deux plaques desmosomales s'appelle la desmoglée. Les desmocollines et desmogléines sont des CAM homophiles (elles entrent en contact avec le même type de molécule), et homotypiques (elles permettent la liaison de deux cellules de même nature). Pour se lier elles ont besoin de Ca2+, qui changent leur conformation et permet de les assembler selon le principe clé-serrure. Les desmosomes sont des dispositifs ponctuels.
On les trouve dans l'épithélium tégumentaire (la surface de la peau), l'endothélium vasculaire. Entre les cellules musculaires et les tendons, il existe des jonctions appelées jonctions myotendineuses qui ont une structure similaire aux desmosomes.
Ils sont composés d'un ensemble de protéines, dont la desmocolline (DSC2), la desmogléine (DSG2), la desmoplakine (DSP), la plakoglobine (JUP), et la plakophiline.
Les hémidesmosomes
Les hémidesmosomes sont semblables aux desmosomes de type macula adherens sauf qu'ils relient la cellule à la lame basale. Il n'y a donc qu'une seule moitié du dispositif en place. Les composants transmembranaires sont l'intégrine alpha6beta4, l'antigène de la pemphigoïde bulleuse de 180kDa ou BP180 et la molécule CD151. La plaque cytoplasmique interne est reliée aux filaments intermédiaires de type cytokératines par l'intermédiaire de la plectine et de l'antigène de la pemphigoïde bulleuse de 230 kDa ou BP230. La laminine est l'intermédiaire entre les différents types de collagène formant la lame basale ou la matrice extracellulaire, et l'hémidesmosomes. NB: les filaments intermédiaires ne se fixent pas de la même façon suivant que ce soit un desmosome ou un hémidesmosome.
Fonctions
Les desmosomes et les hémidesmosomes sont des jonctions qui assurent la solidité mécanique du tissu partout où cela est nécessaire. Un desmosome retient fermement les cellules entre elles de façon qu'elles forment un tissu résistant, renforcé par des filaments intermédiaires. Elles permettent aussi la transmission des signaux intracellulaires, notamment par le biais de la caténine bêta.
Lors de l'embryogénèse, les macula adherens interviennent dans les processus fondamentaux au moment de la mise en place des feuillets embryonnaires et lors de la différenciation du tube neural.
Certaines pathologies sont liées à un dysfonctionnement des desmosomes. En cas de cancer, l'expression des cadhérines des desmosomes ceinturant est diminuée. La mutation des gènes liés aux desmosomes entraine souvent des pathologies cardiaques car les stries scalariformes, très présentes dans le muscle cardiaque strié, sont maintenues conjointes par de nombreux desmosomes.
En médecine
La dysplasie ventriculaire droite arythmogène est une maladie du desmosome cardiaque[1].
Notes et références
- Corrado D, Link MS, Calkins H, Arrhythmogenic right ventricular cardiomyopathy, N Engl J Med, 2017;376:61-72
Voir aussi
Bibliographie
- Bornslaeger, EA, CM Corcoran, TS Stappenbeck, KJ Green (1996) Breaking the connection: displacement of the desmosomal plaque protein desmoplakin from cell–cell interfaces disrupts anchorage of intermediate filament bundles and alters intercellular junction assembly. J Cell Biol 134:985–10018769422 (résumé)
- Boyer, B, GC Tucker, AM Vallés, WW Franke, JP Thiery (1989) Rearrangements of desmosomal and cytoskeletal proteins during the transition from epithelial to fibroblastoid organization in cultured rat bladder carcinoma cells. J Cell Biol 109:1495–15092677020 (résumé)
- Demlehner, MP, S Schafer, C Grund, WW Franke (1995) Continual assembly of half-desmosomal structures in the absence of cell contacts and their frustrated endocytosis : a coordinated Sisyphus cycle. J Cell Biol 131:745–7607593194
- Garrod D & Chidgey M (2008) Desmosome structure, composition and function. Biochimica et Biophysica Acta (BBA)-Biomembranes, 1778(3), 572-587.
- Garrod, DR (1993) Desmosomes and hemidesmosomes. Curr Opin Cell Biol 5:30–408448028
- Hatzfeld M, GI Kristjansson, U Plessmann, K Weber (1994) Band 6 protein, a major constituent of desmosomes from stratified epithelia, is a novel member of the armadillo multigene family. J Cell Sci 107:2259–22707527055 (résumé)
- Koch, PJ, WW Franke (1994) Desmosomal cadherins: another growing multigene family of adhesion molecules. Curr Opin Cell Biol 6:682–6877833048
- Kouklis, PD, E Hutton, E Fuchs (1994) Making a connection: direct binding between keratin intermediate filaments and desmosomal proteins. J Cell Biol 127:1049–10607525601 (résumé)
- Savagner, P., Yamada, K. M., & Thiery, J. P. (1997). The zinc-finger protein slug causes desmosome dissociation, an initial and necessary step for growth factor–induced epithelial–mesenchymal transition. The Journal of cell biology, 137(6), 1403-1419.
- Stappenbeck, TS, EA Bornslaeger, CM Corcoran, HH Luu, ML Virata, KJ Green (1993) Functional analysis of desmoplakin domains: specification of the interaction with keratin versus vimentin intermediate filament networks. J Cell Biol 123:691–7057693716 (résumé)
- Stappenbeck, TS, JA Lamb, CM Corcoran, KJ Green (1994) Phosphorylation of the desmoplakin COOH terminus negatively regulates its interaction with keratin intermediate filament networks. J Biol Chem 269:29351–293547525582 (résumé)
- Troyanovsky, SM, LG Eshkind, RB Troyanovsky, RE Leube, WW Franke (1993) Contributions of cytoplasmic domains of desmosomal cadherins to desmosome assembly and intermediate filament anchorage. Cell 72:561–5747679953
- Troyanovsky, SM, RB Troyanovsky, LG Eshkind, VA Krutovskikh, RE Leube, WW Franke (1994) Identification of the plakoglobinbinding domain in desmoglein and its role in plaque assembly and intermediate filament anchorage. J Cell Biol 127:151–1607929560 (résumé)
- Troyanovsky, SM, RB Troyanovsky, LG Eshkind, RE Leube, WW Franke (1994) Identification of amino acid sequence motifs in desmocollin, a desmosomal glycoprotein, that are required for plakoglobin binding and plaque formation. Proc Natl Acad Sci USA 91:10790–107947971964 (résumé)