Régulation sociale du taux de reproduction
La sociabilité représente les interactions de type social au sein d'un groupe d'individus appartenant à la même espèce et organisé sur un mode coopératif. La coopération présente une grande variabilité de complexité croissante et peut être de types diverses qui vont définir les différents degrés de sociabilité.
L'étude de la régulation sociale des traits d'histoire de vie (THV) permettant de définir les interactions directes entre individus influent sur la variabilité de ces traits d'histoire de vie.
Les THV sont les caractéristiques d'un organisme (d'un génotype) qui ont un effet sur sa valeur sélective (ou fitness). Certains THV sont dits "fondamentaux" car ils sont importants pour caractériser l'espèce étudiée.
Le taux de reproduction est individuel et peut être défini comme le nombre de descendants produits par un même individu. Il correspond à un paramètre qui régule la dynamique des populations. En ce sens, c'est un THV fondamental à ne pas négliger pour de bonnes prédictions écologiques au niveau de la population.
Il y a deux types de régulation sociale du taux de reproduction: la coopération ou la compétition. Ces deux interactions sont importantes à étudier car même dans une société animale, donc un groupement coopératif, il y a mise en place de mécanismes qui tendent à diminuer également le taux de reproduction d'autres individus. Il est important de distinguer la coopération de l'altruisme de reproduction. Dans une société animale on n'observe pas toujours d'altruisme de reproduction (type compétition reproductive) mais l'altruisme de reproduction ne peut s'observer que dans une société animale (degré important de coopération)[1].
Les différents types de sociétés
Il existe différents degrés de sociabilité en fonction du type de comportement coopératif. Cette liste va du degré le plus faible au degré le plus fort :
- Société de type regroupement ou grégaire: Il s'agit d'un groupement d'organismes résultant de facteurs émanant de leurs congénères et non de l'environnement. C'est une forme très simple de la société animale et se caractérise par une simple inter-attraction. Les relations inter-individuelles observées se limitent à une synchronisation des activités de groupes.
- Société de type subsociale: on observe la présence de comportements parentaux (soins aux jeunes).
- Société de type coloniale : (ou regroupement multifamilial) II s'agit de groupements représentant des familles (liens apparentés) mais séparés (pas d'interactions entre familles). Il existe pourtant des sites d'élevages communs pour le soin aux jeunes mais pas de coopération entre soigneurs.
- Société de type communale : Les soigneurs (femelles) coopèrent pour le soin des jeunes.
- Société de type eusociale : On observe: Une coopération dans le soin des jeunes, le chevauchement d'au moins deux générations d'individus et l'existence de castes (individus spécialisés dans la reproduction, avec une division du travail (polyéthisme)). Voici les trois critères nécessaires pour parler d'eusocialité selon Edward O.Wilson.
Il est important de distinguer ces différents types de sociétés car selon les modèles choisis la régulation sociale ne sera pas la même. Certains mécanismes ne sont observés que dans les sociétés de type eusociale et d'autres que dans les sociétés aux degrés de sociabilisation plus faibles.
Compétition et Coopération
- Compétition Il existe plusieurs types de compétitions intraspécifiques, mais celle qui peut être un facteur de régulation sur les taux de reproduction est la contest competition ou compétition par concours qui fait intervenir une interaction directe entre deux individus d'une population pour une même ressource (alimentaire, eau, espace, accès aux partenaires sexuels). La compétition peut entraîner une diminution des ressources, ce qui peut influencer le taux de survie de la population et donc le taux de reproduction. C'est un facteur limitant : Le taux de reproduction est un trait d'histoire de vie dépendant des autres traits. Si l'allocation des ressources diminue, la proportion de ces ressources allouées à la reproduction diminue aussi (effort reproducteur). De plus si les ressources diminuent le taux de survie diminue, la population décroît. Ainsi le nombre d'individus engendrés à la génération suivante dans une population aura tendance à être réduit.
- Coopération Plusieurs types de coopération existent dans différents types de populations et de sociétés. La coopération intraspécifique a pour but d'augmenter la fitness individuelle et donc la densité de la population. La coopération peut ainsi créer de la densité-dépendance positive au sein d'une population. La coopération entre individus implique des coûts. Selon la théorie de l'évolution de la coopération, basée sur le concept de la fitness inclusive , il est largement admis que la sélection peut opérer par le biais de la coopération. Cette coopération peut se faire entre apparentés comme les soins apportés par les parents, (voir théorie de la sélection de parentèle) ou entre non-apparentés. Les mécanismes évolutifs qui maintiennent ce type de relation entre non apparentés reste un sujet de débat. On suppose que les individus qui échangent une ressource ou un service ont plus a y gagner : certes, le service ou la ressource donné(e) à un autre individu demande un coût pour le donneur mais ce coût est vite compensé lorsqu'il devient receveur. On appelle ce mécanisme, la réciprocité directe. Dans certains cas, la coopération génère une synergie immédiate dont les avantages partagés par les coopérateurs dépassent les coûts de l'assistance. Ce type de coopération est retrouvé chez de nombreuses espèces et permet d'augmenter le taux de reproduction individuel au sein de la population. En fonction du niveau de hiérarchisation de la population et des relations dominants-subordonnés, des écarts (variance) de ce taux peuvent être observés : La coopération peut alors prendre la forme de manipulation, dans laquelle l'individu dominant force le ou les subordonnés à être coopératif(s) dans le but d'augmenter sa propre fitness (et donc son taux de reproduction). Cette stratégie de manipulation est en quelque sorte un comportement égoïste et est appelé la tactique coercitive. La coopération au sein d'une population peut alors prendre différentes formes : Le mutualisme, la réciprocité et la manipulation.
- Le mutualisme : Des études récentes ont montré que la coopération peut générer des avantages partagés immédiats. Dans certaines sociétés animales le mutualisme permet de conserver des relations affiliatives à long terme entre individus d'une même population. Chez certains groupes de primates, les individus forment des relations durables entre non apparentés, en pratiquant par exemple l'épouillage mutuel. Selon une étude, les différences d'étendue et de qualité de ces relations peuvent affecter le taux de reproduction à long terme.
Mécanismes sociaux : Les modèles de base
Théorie de la sélection de groupe
Jusque dans le milieu des années 1960, les scientifiques pensaient que les comportements animaux et leurs interactions sociales étaient destinés à assurer la survie de l'espèce et donc du groupe. Le père de cette idée est Wynne-Edwards[2] - [3] qui formule la théorie de la sélection de groupe en 1962. Cette théorie nous dit que les individus seront sélectionnés pour sacrifier leur propre intérêt (diminuer sa fitness individuelle) au profit du groupe, de l'espèce (fitness globale). Ce « sacrifice » permettrait de réguler l'utilisation des ressources de manière à éviter une pénurie qui mènerait à l'extinction de l'espèce (ou à une autre échelle, du groupe). Ainsi il y aurait mise en place d'une forme de coopération où tous les individus régulent leur taux de reproduction (contrôle des naissances, etc.) pour préserver l'environnement et les ressources liés à celui-ci. Les individus incapables d'effectuer ce mécanisme d'auto-régulation seraient menacés d'extinction par surexploitation de leur milieu[1].
On parle bien d'ici d'une régulation sociale (groupe, coopération...) sur le taux de reproduction.
Théorie de la sélection de parentèle
Cette théorie de la sélection de la parentèle a été développée par William Donald Hamilton dans les années 1960 pour expliquer l'observation de l’altruisme de reproduction dans les populations. Cette théorie démontre notamment que la sélection de groupe n'explique pas de manière fiable l'apparition de la sociabilité. La base de cette théorie est que l'altruisme au sein d'une population s'observera lorsque les coûts (fitness réduite) dus à la coopération sont inférieurs aux bénéfices (fitness augmentée) pour l'individu aidé. De plus cette relation est d'autant plus vraie si on prend en compte le degré d'apparentement entre les deux individus ce qui augmente d'autant plus les bénéfices dus à la coopération.

c = coûts pour l'individu qui coopère.
R= coefficient d'apparentement entre individu qui coopère et individu receveur.
b= bénéfices pour l'individu qui reçoit.
La sélection naturelle sélectionne donc ce comportement lorsque la coopération se fait entre individus apparentés :
Valeur sélective directe + VS indirecte = VS totale.
Où VS directe = succès reproducteur propre de l'individu et où VS indirecte = l'aide liée au succès reproducteur à un individu apparenté.Si on prend par exemple le comportement d'assistance des jeunes vers leurs parents pour élever ses frères et sœurs, le taux de reproduction du jeune assistant se verra diminué au profit d'une fitness augmentée car la transmission de ses gènes à la génération suivante est de 0,5.
.pdf.jpg.webp)
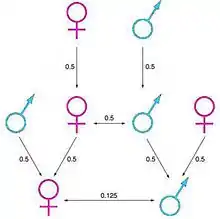
Mécanismes sociaux : Régulations sur les taux de reproduction
Effet Allee
L'effet Allee est un mécanisme clé dans la régulation des dynamiques des populations. Il y a une corrélation positive entre taux de croissance individuel dans une population quand la population est de petite taille. Les effets Allee peuvent être classés en deux groupes : les effets forts et les effets faibles:
- Les effets Allee forts sont caractérisés par un seuil, représentant la taille de la population critique, en dessous duquel le taux de croissance par individu devient négatif.
- Un effet Allee est dit faible, lorsque la relation taux de croissance d'un individu et croissance démographique reste positive.
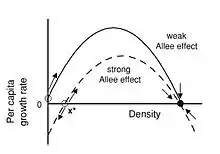
L'effet Allee influe sur la probabilité d'extinction d'une population de faible densité. Il est donc important de déterminer les taux de reproduction et les taux de croissance afin d'établir les prédictions les plus précises. Démontrer l'effet Allee de façon expérimentale devient rapidement difficile voire impossible et diffère selon les espèces. Dans les populations où les organismes ont une taille réduite ou à vie fixée, l'effet Allée peut être démontré sur le terrain ou en laboratoire. Mais cela est généralement impossible dans les populations sauvages dont les individus possèdent une grande taille (ex: les Mammifères). Néanmoins, ce n'est pas parce qu'un effet Allee n'est pas observable qu'il peut être ignoré dans l'étude de la viabilité d'une espèce, donnée par la relation suivante :

Avec N, la densité de la population; K, la capacité de charge du milieu; A, la densité critique de la population.
Un effet Allee est dit composant, lorsqu'il y a une relation positive entre tout composant de la fitness individuelle (taille, densité, conditions physiques) et traits démographiques (taux de reproduction, de croissance…). Nous pouvons observer des effets Allee lorsqu'il n'y a pas de relations de compétition au sein de la population.Un effet Allee est dit démographique lorsqu'il se réfère à une densité-dépendance positive sur le taux de croissance d'un individu au sein d'une population. Le modèle de l'effet Allee doit être lié à un modèle de dynamique des populations, dans lequel l'effet Allee composant peut se traduire par un effet démographique (ex : Variation du taux de reproduction). Par exemple chez les suricates, la relation entre individu dominant-dominé permet d'augmenter la densité-dépendante positive (et donc de provoquer un effet Allee): seule la femelle dominante peut se reproduire. Les subordonnés non reproducteurs, n'ayant pas de progéniture vont pouvoir accorder tous les soins nécessaire à la femelle et sa future descendance. Ainsi, le taux de survie des jeunes viables est augmenté. Le taux de reproduction chez la femelle dominante augmente pour ce type de coopération mais est nulle pour les autres individus. Il serait donc intéressant de calculer la variance dans ce type de population où la hiérarchisation occupe une place importante[4] - [5] - [6] - [7].
Suppression de reproduction
La suppression de reproduction (en) peut être définie comme une inhibition de deux paramètres : les mécanismes physiologiques de reproduction et/ou les comportements reproducteurs chez un individu fertile. Via cette définition, la suppression de reproduction est un changement dans le processus normal de reproduction de l'individu par une influence inhibitrice. Ce changement peut être observé dans de nombreux taxons différents et chez les deux sexes. L'influence inhibitrice peut être de différentes natures, une pression interne ou externe (via l'environnement) et peut intervenir dans les différents stades du processus reproductif.
Le processus reproductif peut être extrêmement coûteux sur le point de vue énergétique et augmente les risques de mortalité, ce qui représente des contraintes encore plus grandes lorsque l'on sait que la probabilité de survie des jeunes produits dépend de nombreux facteurs : conditions physiques, conditions biotiques de l'environnement, interactions sociales au sein de la population...
Une suppression de reproduction connue et observable facilement est le fait de réguler sa reproduction annuelle avec la mise en place de périodes de reproduction. La reproduction étant coûteuse et risquée, les individus se reproduisent seulement au moment de l'année qui réunit les meilleures conditions pour « rentabiliser » la reproduction. La sélection naturelle tend à avantager la régulation de la reproduction via la suppression pour optimiser ce processus coûteux.
La suppression de reproduction peut être expliquée par deux influences : limites physiologiques et limites externes.
La première est souvent observée chez les Mammifères qui, en période de faibles ressources (ou faibles températures... Facteurs biotiques changés), entrent dans un état où la balance énergétique est faible. Il y a mise en place d'une signalisation métabolique qui déclenche ou non le processus reproducteur en fonction de l'état physiologique de l'individu. Cette signalisation est encore mal connue aujourd'hui. La physiologie influence la reproduction également si l'individu est malade.
Les facteurs externes (généraux) pouvant supprimer (ou activer) la reproduction sont l'alimentation, les facteurs physiques ou l'influence sociale.
Suppression de reproduction pré-conception
Nous allons nous intéresser ici à l'influence sociale sur la suppression de reproduction.
L'environnement social peut en effet supprimer en partie ou totalement le processus physiologique et/ou comportemental de la reproduction, chez beaucoup d'espèces. Cette influence sociale provient souvent des relations agonistes ou dominance/dominé entre des individus adultes du même sexe.
Par exemple, chez les rats taupes (Heterocephalus Glaber) et chez les ouistitis (Callithrix jacchus) on observe une suppression de reproduction chez certaines femelles. En présence de la femelle dominante (femelle principale pour la reproduction), les femelles subordonnées ont un taux faible d'hormones de reproduction, n'ovulent pas ou peu et n'ont pas un comportement sexuel observable. Cette fonction de reproduction peut réapparaître s'il y a éloignement des femelles subordonnées vis-à-vis de la femelle dominante. Il y a bien suppression de reproduction par une pression sociale de type dominant/dominés.
Les interactions dominant/dominés ne sont pas le seul processus social qui influence la reproduction. Chez les souris domestiques (Mus musculus) lorsque la densité de la population augmente il y a production de signaux chimiques qui régulent l'ensemble des cycles œstraux pour réduire la fertilité des femelles (densité-dépendance).
Cette suppression de reproduction est moins fréquente chez les mâles que chez les femelles, cela est dû au coût plus faible chez les mâles pour la reproduction. Les deux types de suppressions observables chez les mâles sont :
- un retard dans la maturité sexuelle en présence d'un mâle dominant (observable chez les Orangs-outans par exemple)
- une inhibition temporaire de l'activité reproductive en présence d'un mâle dominant (observable chez les Marmottes, les Rats taupes, etc.).
La suppression de reproduction engendrée par une pression sociale peut être également expliquée par une simple réponse par rapport au stress engendré par les interactions entre individus.
Suppression de reproduction post-conception
La régulation sociale sur la reproduction peut se faire à plusieurs niveaux de la reproduction. On a vu précédemment que la régulation peut se faire en inhibant la reproduction en amont (retard de la maturité par exemple), mais elle peut se faire après cette reproduction. Lorsque les femelles dominantes ne peuvent pas inhiber la conception des femelles subordonnées, elles passent par des voies alternatives comme l'avortement, les expulsions des subordonnées ou l'infanticide[8] - [9].
Exemple de l'infanticide: ce qui désigne le meurtre d'un descendant. L'infanticide peut être une régulation majeure dans une structure sociale lorsque les femelles entretiennent une longue relation avec les mâles.

Trois conditions (dictées par la théorie de la sélection sexuelle) pour observer un infanticide : le mâle n'a pas ou peu de probabilité d'être le père de l'enfant. La femelle pourra être capable de se reproduire rapidement après l'infanticide. Le mâle augmente sa probabilité de s'accoupler et d'être le père des futurs descendants de cette femelle. C'est le contexte dans lequel on peut observer de l'infanticide dans une sociétés dominant/dominés (exemple des Gorilles de montagne)[10].
L'infanticide permet aussi d'imposer la période de reproduction de certains individus comme chez les fourmis où les ouvrières tuent tous les œufs mâles durant toute l'année sauf pendant la période de reproduction. Ici c'est le taux de reproduction de la population qui est régulé socialement[11].
La hiérarchisation
La régulation sociale permet de maintenir une certaine structure et organisation des groupes sociaux. Cette régulation se base sur la hiérarchisation au sein des espèces où chaque individu doit trouver sa place, ses fonctions par rapport aux autres.
Mise en place de la hiérarchie au sein d’un groupe.
Mécanisme de compétition pour un territoire :
Chez certaines espèces dès le plus jeune âge, la mise en place d’un territoire s’avère cruciale pour leur place au sein de la communauté. L’acquisition d’un territoire est déterminée par des interactions biotiques. Elle se réalise par la capacité des jeunes à coloniser leur environnement mais également par les interactions entre les individus pour la défense de cet espace acquis. Cela représente une adaptation comportementale par l’exploitation de ressources et tend vers un ajustement de la densité de l’espèce, également ajustée par le taux de mortalité des jeunes.
La compétition pour un territoire induit des interactions fortes entre individus. La défense de celui-ci amène à une gamme de comportements agonistiques qui va de la tendance la plus agressive à la soumission.L’individu définit alors sa place hiérarchique qui induit la stabilité du groupe. Un plus grand territoire apporte une place hiérarchique plus élevée ainsi qu’un avantage compétitif. Un espace plus grand permet à l’individu un meilleur taux de croissance, une meilleure survie ainsi qu’une meilleure reproduction.
Statut de reproducteur
Au sein de certaines espèces il peut y avoir des conflits au niveau de la reproduction. La régulation de ce partage pour la reproduction se ferait grâce à la détection d’hormones de fertilités et permettrait aux individus d’ajuster leur comportement reproducteur selon le contexte social et leur intérêt du point de vue du fitness. Une hiérarchisation se met donc en place avec le plus souvent un seul individu ou un couple d’individus pouvant se reproduire. Leur taux de reproduction est augmenté et celui des autres diminué ou complètement inhibé. Si l’individu meurt la hiérarchisation reproductive linéaire permet aux individus du rang suivant d’accéder à la reproduction. Ce statut reproducteur joue donc un rôle important dans les interactions de dominance/subordination qui sont la base de la hiérarchie sociale[12].
Les limites d'un modèle fixe pour ce sujet d'étude
Les mécanismes généraux de régulation sociale du taux de reproduction ont été cités dans les parties précédentes de cette page. Mais lorsque l'on commence à choisir un modèle d'étude on observe l'apparition de mécanismes spécifiques au modèle. En effet on peut observer diverses nuances:
- Différences entre les degrés de sociabilisation.
- Différences entre sociétés du même degré de sociabilisation.
- Différences selon la période d'étude[10].
Effets de taille, biais reproducteur et complexité sociale
Les espèces peuvent être placées sur un « continuum » selon leur degré de complexité social.
La taille des groupes sociaux est un facteur important de ce continuum car elle agit sur la tendance à se reproduire des assistants et sur la nature des conflits de manière similaire chez de nombreuses espèces dans provenant de taxons différents[13].
La transition des sociétés simples aux sociétés complexes est corrélée positivement avec le biais de reproduction et le nombre d’effectifs. Ainsi, les sociétés les plus complexes sont celles des insectes qui réunissent une femelle reproductrice qui dépend du plus grand nombre de membres non-reproducteurs et la plus grande variation morphologique (correspondant à une haute spécialisation des membres)[14], alors que les plus simples sont caractérisées par un élevage des jeunes principalement effectué par les parents. Dans cette dernière, qualifiée de « subsocial », la compétition pour les ressources augmente et le succès reproducteur diminue ou reste constant avec une augmentation de la taille du groupe. À l’intermédiaire, se placent les groupes qui partagent les soins des jeunes en communauté et qui présentent une relation positive entre la taille du groupe et le succès reproducteur (« communal »)[15].
Effets de la composition du groupe sur la reproduction
Selon la théorie de la sélection de parentèle, les animaux vont préférentiellement se regrouper entre individus de proche parenté pour augmenter leur valeur sélective totale en aidant un individu apparenté. De plus, via une augmentation de la tolérance entre individus et une diminution des risques d’infanticides, les groupes sociaux composés de membres apparentés devraient avoir un taux de reproduction plus fort que chez les groupes sociaux composés d’individus non apparentés[15] - [16]
Exemple : Chez de nombreux rongeurs Arvicolinae, la densité de la population suit un cycle de 3-5 ans et cette dynamique a été partiellement expliquée par la préférence des individus à se regrouper entre membres apparentés. À la suite de l’augmentation de la densité de la population et les limites fixées par les territoires, un phénomène de dispersion va avoir lieu sur de grandes distances. Les nouveaux territoires avoisinants, composés d’individus plus distants génétiquement, vont voir leur densité de population diminuer à la suite de l’augmentation des agressions et de la suppression de la reproduction[15] - [17].
Mais il existe des contre-exemples comme:
Boonstra & Hogg (1988) ont comparé un enclos de campagnols composé de femelles apparentées (3 sœurs + filles descendantes) avec celui composé de femelles non apparentées. Même si les femelles apparentées avaient une taille de portée légèrement supérieure, au bout d’une période de 6 mois, les deux populations étaient de densité et de taille équivalente[17].
Le mécanisme "général" de la sélection de parentèle explique dans beaucoup d'espèces l'apparition de régulation sociale sur le taux de reproduction mais, selon les modèles, d'autres mécanismes interviennent.
L'intégration sociale sur le succès reproducteur des femelles

L’intégration sociale des femelles qui vivent au sein de groupes sociaux stables et à long-terme peut permettre une augmentation de la survie des jeunes et un meilleur taux de reproduction, indépendamment de la qualité de l’habitat, la composition du groupe social, du statut de dominance et de l’âge. Cette intégration sociale a un bénéfice direct sur le fitness en réduisant les harcèlements qui sont source de stress, et/ou se protéger contre les risques d’infanticides.
Par exemple : chez les équidés, les femelles qui ne sont pas loyales à un groupe de reproduction et qui négligent les interactions sociales comme le grooming (épouillage/toilettage), souffrent d’agressions répétitives et sont relativement en mauvaise santé avec un succès reproducteur faible[18].
L’importance de l’intégration sociale est apparente lorsqu'une forte intégration peut compenser la disparité du succès reproducteur liée à une position de bas rang d’un individu subordonné.
Une femelle qui « groom » de façon équitable et qui a les relations sociales les plus robustes et les plus durables pourrait atteindre les mêmes taux de reproduction que la femelle dominante[15] - [19].
Différences entre modèles de type eusocial
Il existe une ressemblance forte entre les petites colonies d’insectes sociaux avec les groupes sociaux des vertébrés, cependant les sociétés des vertébrés n’atteignent jamais le même degré de complexité[14] - [20].
Seules deux espèces de mammifères s’y approchent en formant des colonies eusociales de 70 à 300 individus avec une seule femelle qui monopolise la reproduction, le rat-taupe glabre et le rat-taupe de Damaraland .

Le rat-taupe nu est particulier au sein des mammifères car sa tendance incestueuse est très élevée. La relation de parentèle fut proposée pour expliquer son eusocialité mais il s’avère que ces colonies suivent le schéma classique de « l’incest avoidance » ("éviter l'inceste") et ceci est suffisant pour maintenir le biais reproducteur si les seuls individus non-apparentés sont les parents fondateurs[21]. Ce mécanisme de contrôle sur la reproduction est si efficace que la mort de la reine peut induire une quiescence de la colonie de plusieurs années jusqu’à l’arrivée d’un individu non-apparenté[21] - [22].
Cependant, le outbreeding (exogamie), qui favorise le brassage génétique est préféré mais la reine est capable de réprimer physiologiquement la reproduction des deux sexes de manière réversible selon le modèle du contrôle par le dominant (« dominant control model »)[21] - [23].
Chez les insectes eusociaux, la transition entre les sociétés simples et complexes est associée à une perte de la capacité des individus à effectuer à la fois un rôle de reproducteur et un rôle d’assistanat/ouvrières de manière irréversible, d’un fort biais reproducteur et une augmentation de la taille des colonies[14] - [24].
Les petites colonies sont alors caractérisées par un faible biais reproducteur, un fort potentiel reproducteur adulte, un degré de dimorphisme entre les rôles entrepris relativement faible et un court cycle de vie de la colonie, alors que les grandes colonies ont un fort biais reproducteur[20], un faible potentiel reproducteur adulte, des ouvrières qui peuvent être polymorphiques (chez les fourmis et les termites) et qui effectuent une inhibition mutuelle de la reproduction (worker policing), et des colonies pérennes avec des relations de parentèle relativement stables[14].
Au sein des colonies de petites tailles, le remplacement de la reine et la reproduction des ouvrières sont fortement sélectionnés. Le conflit entre reines et ouvrières pour la reproduction peut s’exercer de manière physique (compétition directe). Effectivement, le faible dimorphisme et le peu de concurrents permettent aux ouvrières de défier la reine et cette dernière doit constamment maintenir la suppression de la reproduction[14] - [20].
Au sein des colonies de grandes tailles, les ouvrières plus petites sont moins sélectionnées pour le remplacement de la reine ou pour la reproduction, et le dimorphisme important engendre des conflits entre reines et ouvrières qui se déroulent de manière indirecte au niveau de la composition de la génération suivante[14].
On observe également des mécanismes bien particuliers à des espèces de type eusocial comme le parasitisme intra-spécifique chez Bombus terrestris (Bourdon terrestre). Les régulations sociales sur le taux de reproduction peuvent être expliquées généralement par la théorie de la sélection de parentèle. Mais on observe l'intervention douvrières parasites qui vont produire des mâles dans la colonie parasitée. Cette intervention influe sur le taux de reproduction des ouvrières résidentes car il y aura dilemme entre le fait de se reproduire avec un individu apparenté (augmenter sa fitness) ou prendre le risque de se reproduire avec un mâle parasite (diminuer sa fitness)[14].
Discussion: Une histoire de stress ?
Beaucoup de nouvelles recherches cherchant à cerner et à mesurer la régulation sociale du taux de reproduction proposent une nouvelle approche.
Elle consiste à simplifier les changements observés du taux de reproduction face à une interaction sociale comme une simple réponse à un stress engendré par un individu directement. En effet, les limites sont nombreuses pour ce genre d'études. Lorsque l'on tente de mesurer ces phénomènes sur le terrain on peut avoir une bonne vision des différentes interactions qui se font entre individus mais il est difficile de mesurer les variations du taux de reproduction. Au contraire en laboratoire cette mesure peut se faire facilement mais les interactions ne se font pas en conditions naturelles et peuvent donc être modifiées. De plus, il est difficile de quantifier l'effet de la reproduction pré-conception car on observe bien l'interaction (expulsion ou attaque) mais on ne sait pas quel est l'effet sur la physiologie de l'animal. En réduisant cette réponse à du stress, on peut éclaircir les événements. Le modèle simple qui en découle est: l'interaction (souvent violente) entre le dominant et le dominé déclenche une réponse physiologique chez le dominés. Celui-ci va produire des hormones de stress (par exemple une hormone surrénale appelée glucocorticoïde (GC)) . Ces hormones de stress ont la particularité d'inhiber ou de favoriser certaines fonctions physiologiques. En l’occurrence, ici, cette hormone peut inhiber la production d'hormones sexuelles et/ou réduire la sensibilité entre l'hormone et ses récepteurs et/ou inhiber l'action de cette hormone. Ces effets ou ces productions d'hormones sont facilement quantifiables en laboratoire. Et en effet, on observe bien corrélation entre ces actions du stress et l'inhibition de la physiologie de reproduction (augmentation des avortements, impossibilité de se reproduire, gonades réduites....)[25].
Références
- Serge Aron et Luc Passera, Les sociétés animales : évolution de la coopération et organisation sociale, Bruxelles/Paris, De Boeck Supérieur, , 336 p. (ISBN 978-2-8041-2861-6, lire en ligne)
- (en) « Animal Dispersion in Relation to Social Behaviour », Ibis, vol. 104, , p. 570-571 (ISSN 1474-919X, DOI 10.1111/j.1474-919X.1962.tb08690.x, lire en ligne, consulté le )
- (en) Mark V. Flinn, « Evolution Through Group Selection. V. C. Wynne-Edwards. », American Anthropologist, vol. 89, , p. 728-729 (ISSN 1548-1433, DOI 10.1525/aa.1987.89.3.02a00280, lire en ligne, consulté le )
- (en) Tim Clutton-Brock, « Cooperation between non-kin in animal societies », Nature, vol. 462, , p. 51-57 (ISSN 0028-0836, DOI 10.1038/nature08366, lire en ligne, consulté le )
- (en) S. Rosenbaum, A. A. Maldonado-Chaparro et T. S. Stoinski, « Group structure predicts variation in proximity relationships between male-female and male-infant pairs of mountain gorillas (Gorilla beringei beringei) », Primates; Journal of Primatology, (ISSN 1610-7365, PMID 26386711, DOI 10.1007/s10329-015-0490-2, lire en ligne, consulté le )
- (en) M. B. V. Bell, M. A. Cant, C. Borgeaud et N. Thavarajah, « Suppressing subordinate reproduction provides benefits to dominants in cooperative societies of meerkats », Nature Communications, vol. 5, (PMID 25047446, PMCID 4109011, DOI 10.1038/ncomms5499, lire en ligne, consulté le )
- (en) Péter K. Molnár, Mark A. Lewis et Andrew E. Derocher, « Estimating Allee Dynamics before They Can Be Observed: Polar Bears as a Case Study », PLoS ONE, vol. 9, (ISSN 1932-6203, PMID 24427306, PMCID 3888426, DOI 10.1371/journal.pone.0085410, lire en ligne, consulté le )
- (en) Wendy Saltzman, The Encyclopedia of Applied Animal Behaviour and Welfare, Wallingford, CABI Press, (lire en ligne), Reproductive suppression, pp 522-523
- (en) Jason S. Gilchrist, « Female eviction, abortion, and infanticide in banded mongooses (Mungos mungo): implications for social control of reproduction and synchronized parturition », Behavioral Ecology, vol. 17, , p. 664-669 (ISSN 1045-2249 et 1465-7279, DOI 10.1093/beheco/ark012, lire en ligne, consulté le )
- (en) Andrew M. Robbins, Maryke Gray, Augustin Basabose et Prosper Uwingeli, « Impact of Male Infanticide on the Social Structure of Mountain Gorillas », PLoS ONE, vol. 8, (ISSN 1932-6203, PMID 24223143, PMCID 3819382, DOI 10.1371/journal.pone.0078256, lire en ligne, consulté le )
- (en) Serge Aron, Luc Passera et Laurent Keller, « Queen-worker conflict over sex ratio: A comparison of primary and secondary sex ratios in the Argentine ant, Iridomyrmex humilis », Journal of Evolutionary Biology, vol. 7, , p. 403-418 (ISSN 1420-9101, DOI 10.1046/j.1420-9101.1994.7040403.x, lire en ligne, consulté le )
- Jacques Gervet, « La Ponte Et Sa Régulation Dans La Société Polygyne De Polistes Gallicus L », Behaviour, vol. 25, , p. 221-233 (ISSN 1568-539X, DOI 10.1163/156853965X00147, lire en ligne, consulté le )
- (en) Daniel T. Bhumstein et Kenneth B. Armitage, « Life history consequences of social complexity a comparative study of ground-dwelling sciurids », Behavioral Ecology, vol. 9, , p. 8-19 (ISSN 1045-2249 et 1465-7279, DOI 10.1093/beheco/9.1.8, lire en ligne, consulté le )
- (en) Bourke, « Colony size, social complexity and reproductive conflict in social insects », Journal of Evolutionary Biology, vol. 12, , p. 245-257 (DOI 10.1046/j.1420-9101.1999.00028.x, lire en ligne, consulté le )
- (en) Joan B. Silk, « The adaptive value of sociality in mammalian groups », Philosophical Transactions of the Royal Society of London B: Biological Sciences, vol. 362, , p. 539-559 (ISSN 0962-8436 et 1471-2970, PMID 17363359, PMCID 2346516, DOI 10.1098/rstb.2006.1994, lire en ligne, consulté le )
- (en) E. L. Charnov et J. P. Finerty, « Vole population cycles: A case for kin-selection? », Oecologia, vol. 45, , p. 1-2 (ISSN 0029-8549 et 1432-1939, DOI 10.1007/BF00346698, lire en ligne, consulté le )
- (en) Rudy Boonstra et Ian Hogg, « Friends and strangers: a test of the Charnov-Finerty Hypothesis », Oecologia, vol. 77, , p. 95-100 (ISSN 0029-8549 et 1432-1939, DOI 10.1007/BF00380931, lire en ligne, consulté le )
- (en) Elissa Z. Cameron, Trine H. Setsaas et Wayne L. Linklater, « Social bonds between unrelated females increase reproductive success in feral horses », Proceedings of the National Academy of Sciences, vol. 106, , p. 13850-13853 (ISSN 0027-8424 et 1091-6490, PMID 19667179, PMCID 2728983, DOI 10.1073/pnas.0900639106, lire en ligne, consulté le )
- (en) S. P. Henzi, L. Barrett, D. Gaynor et J. Greeff, « Effect of resource competition on the long-term allocation of grooming by female baboons: evaluating Seyfarth's model », Animal Behaviour, vol. 66, , p. 931-938 (DOI 10.1006/anbe.2003.2244, lire en ligne, consulté le )
- (en) Paul W. Sherman, Eileen A. Lacey, Hudson K. Reeve et Laurent Keller, « Forum The eusociality continuum », Behavioral Ecology, vol. 6, , p. 102-108 (ISSN 1045-2249 et 1465-7279, DOI 10.1093/beheco/6.1.102, lire en ligne, consulté le )
- (en) C. G. Faulkes et N. C. Bennett, « Family values: group dynamics and social control of reproduction in African mole-rats », Trends in Ecology & Evolution, vol. 16, , p. 184-190 (ISSN 1872-8383, PMID 11245941, lire en ligne, consulté le )
- (en) J. U. M. Jarvis et N. C. Bennett, « Eusociality has evolved independently in two genera of bathyergid mole-rats — but occurs in no other subterranean mammal », Behavioral Ecology and Sociobiology, vol. 33, , p. 253-260 (ISSN 0340-5443 et 1432-0762, DOI 10.1007/BF02027122, lire en ligne, consulté le )
- (en) « Proximate mechanisms regulating a reproductive dictatorship: a single dominant female controls male and female reproduction in colonies of naked mole-rats », sur ResearchGate (consulté le )
- (en) Bernard J. Crespi et Douglas Yanega, « The definition of eusociality », Behavioral Ecology, vol. 6, , p. 109-115 (ISSN 1045-2249 et 1465-7279, DOI 10.1093/beheco/6.1.109, lire en ligne, consulté le )
- (en) Andrew J. Young, Anne A. Carlson, Steven L. Monfort et Andrew F. Russell, « Stress and the Suppression of Subordinate Reproduction in Cooperatively Breeding Meerkats », Proceedings of the National Academy of Sciences of the United States of America, vol. 103, , p. 12005-12010 (lire en ligne, consulté le )