Effet Allee
L'effet Allee est un phénomène de dépendance positive en la densité en écologie des populations, caractérisé par une corrélation positive entre la densité d'une population et son taux de croissance. Il se manifeste chez certaines espèces et seulement pour de faibles effectifs.
Historique
Le phénomène a été décrit pour la première fois en 1931 par Warder Clyde Allee (1885-1955). Allee était un zoologiste américain qui portait un intérêt particulier au comportement animal. Professeur à l’université de Chicago, il était d’ailleurs l'un des premiers à vouloir établir l’écologie comme science indépendante au sein de la biologie[1].
Si la plupart de ses contemporains se fondaient sur le principe malthusien (croissance exponentielle) et le modèle de Verhulst (croissance logistique), Allee avait constaté que ce n’est pas forcément la compétition intraspécifique qui limite la croissance d’une population. Un faible effectif (en anglais : undercrowding) peut aussi réduire la croissance alors que l’agrégation peut avoir un effet positif. Allee détaille ces observations dans son livre de 1949 Principles of Animal Ecology en énonçant que la coopération intraspécifique conduit à une dépendance en la densité inverse de celle que l'on observe généralement et que donc la croissance per capita est diminuée si la taille de la population est (trop) faible[2].
P.E. Odum, un collaborateur de W.C. Allee, introduit en 1959 le terme de « principe Allee » qui après a été repris sous la forme d'« effet Allee »[2].
Description
La présence d'un effet Allee dans une population a été décrite comme l'existence d'une dépendance en la densité inverse de celle que l'on observe généralement, pour de faibles effectifs. C'est-à-dire l'existence d'une corrélation positive entre la densité d'une population et son taux de croissance lorsque la population comporte relativement peu d'individus[2]. Cela résulte en une diminution du taux de croissance de la population quand sa taille diminue, et inversement une augmentation de ce taux quand sa taille augmente. Ce principe s'oppose à l'idée qu'il existe toujours une corrélation négative entre la densité d'une population et son taux de croissance, liée essentiellement aux phénomènes de compétition. De très nombreux mécanismes peuvent donner naissance à un effet Allee dans une population à faible densité, comme la consanguinité, la stochasticité démographique et les phénomènes de facilitation[2].
On distingue deux formes de l'effet Allee : l'effet Allee élémentaire (component Allee effect) et l'effet Allee démographique[3] - [4].
- Un effet Allee élémentaire est observé quand un composant de la valeur sélective individuelle, par exemple le taux de survie ou de reproduction, est positivement corrélé à la taille de la population. Ainsi, dans une même population il peut y avoir de nombreux effets Allee élémentaires.
- Un effet Allee démographique, en revanche, ne se manifeste que si les effets Allee élémentaires ne sont pas contrebalancés par des phénomènes de dépendance négative en la densité, comme la compétition. Dans ce cas, l'effet Allee s'observe à l'échelle de la population et influence la dynamique de cette dernière, pour des faibles densités.
À noter que pour constater un effet Allee démographique, il suffit d'observer l'évolution de la population à des faibles densités, alors que pour un effet Allee élémentaire, il faut parvenir à mesurer la variation de composants particuliers de la valeur sélective à des faibles densités de population, ce qui est plus difficile à réaliser. Par conséquent, bien que théoriquement admis depuis longtemps, l'effet Allee élémentaire a été peu observé dans des populations naturelles, contrairement à l'effet Allee démographique dont on a trouvé des manifestations dans la majorité des grands taxa animaux. Par la suite on désignera donc par effet Allee l'effet Allee démographique.
On sépare deux intensités d'effet Allee : fort ou faible[3].
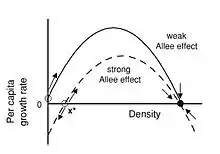
- L'effet Allee est considéré comme fort lorsqu'il existe une densité de population, dite critique, en dessous de laquelle le taux de croissance par individu est négatif.
- L'effet Allee est dit faible si le taux de croissance par individu est toujours positif, mais plus faible pour des faibles densités que pour des densités plus élevées.
Quoique difficile à observer dans des populations naturelles, l'effet Allee est un principe théoriquement simple qui peut aisément être observé dans des modèles simples de dynamique de population.
L'effet Allee dans le modèle logistique
Mise en équation
Le modèle de Verhulst est un modèle simple de dynamique de population qui considère la diminution du taux de croissance de la population avec l'augmentation de sa taille (dépendance négative en la densité liée à la compétition) et une capacité de charge du milieu, définie comme le nombre d'individus maximal de cette population que peut supporter le milieu, en posant l'équation suivante :

où :
- N(t) correspond à l'effectif de la population et varie au cours du temps.
- r est une constante correspondant au taux de croissance intrinsèque de la population.
- K est une constante correspondant à la capacité de charge du milieu.
- dN(t)/dt correspond à la variation de l'effectif au cours du temps, c'est-à-dire au taux de croissance moyen de la population.
Dans ce modèle, on peut rajouter un terme d'effet Allee, ce qui donne :

où :
- A correspond à la densité critique en dessous de laquelle le taux de croissance de la population devient négatif.
Remarque : Pour un effet Allee faible, il n'y a pas de densité critique A et l'équation peut s'écrire :

Remarque : K est aussi bien une caractéristique du milieu que de la population, car de nombreux caractères peuvent influencer la façon dont les individus exploitent les ressources. A est également une caractéristique de la population. Pour une population naturelle, ces deux constantes sont obtenues expérimentalement en observant la dynamique de population.
On peut également diviser cette équation par N(t) des deux côtés pour obtenir :

où :
- dN(t)/dt*1/N(t) est le taux de croissance per capita (par individu). On raisonnera sur cette deuxième équation pour la suite.
Équilibres
On peut alors s'intéresser aux équilibres de cette dynamique de population, c'est-à-dire aux points où le taux de croissance per capita est nul. Graphiquement parlant, sur la courbe du taux de croissance per capita en fonction de l'effectif de la population, les équilibres sont les points où la courbe coupe l'axe des abscisses.
Pour un effet Allee fort :
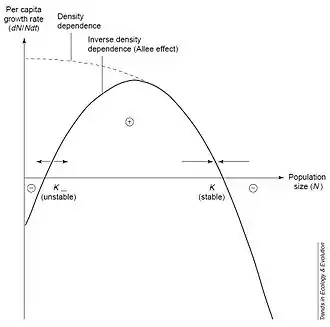
On observe deux équilibres :
- Un pour un effectif A : N(t) = A. Cet équilibre est prévisible, puisque par définition de l'effet Allee, A est l'effectif critique en dessous duquel le taux de croissance est négatif.
- Un pour un effectif K : N(t) = K. Cet équilibre est également prévisible puisque K est l'effectif maximal d'individus de la population que peut supporter le milieu.
Stabilité de ces équilibres :
Ces équilibres peuvent être stables ou instables :
- Dans le cas présent, un équilibre est stable si une petite variation d'effectif autour de l'équilibre est compensée par la variation du taux de croissance. C'est-à-dire :
- À l'équilibre, par définition le taux de croissance est nul.
- Quand on augmente un peu l'effectif, le taux de croissance devient négatif et ramène l'effectif à l'équilibre.
- Quand on diminue un peu l'effectif, le taux de croissance devient positif et ramène l'effectif à l'équilibre.
- Inversement, l'équilibre est instable si une petite variation de l'effectif autour de l'équilibre n'est pas compensée, mais amplifiée par la variation du taux de croissance :
- Quand on augmente un peu l'effectif, le taux de croissance devient positif, augmentant encore davantage l'effectif.
- Quand on diminue un peu l'effectif, le taux de croissance devient négatif, diminuant encore davantage l'effectif.
Il en résulte que pour un effet Allee fort, l'équilibre à N(t) = A est instable et l'équilibre à N(t) = K est stable[2].
Que l'équilibre soit stable à K, la capacité de charge du milieu, est une caractéristique du modèle logistique. En revanche, l'instabilité de l'équilibre à A, l'effectif critique, est ce qui fait tout l'intérêt de l'effet Allee, en particulier dans des problématiques de conservation de populations à petits effectifs. Tant qu'on reste au-dessus de A la population ne devrait pas s'éteindre. En revanche si on passe en dessous de A, le taux de croissance devient négatif, causant une diminution des effectifs, qui elle-même cause une diminution encore plus grande du taux de croissance et ainsi de suite, ce qui peut causer une extinction très rapide de la population. Pour cette raison, la détermination de A dans les populations à protéger ou à réinsérer est un point important et a donné lieu à de nombreuses études[3].
Remarques :
Pour un effet Allee faible, par définition, A n'existe pas et donc on observe un équilibre instable en 0 et un équilibre stable en K. La population ne risque donc pas de spirale d'extinction à de faibles densités, mais souffre cependant de plus faibles taux de croissance que s'il n'y avait aucun effet Allee. À de fortes densités, dans la plupart des cas la compétition prend le pas sur l'effet Allee, et le taux de croissance varie comme pour une population sans effet Allee : selon une dépendance négative en la densité (croissance logistique simple).
Mécanismes
Il existe plusieurs mécanismes qui peuvent induire un effet Allee élémentaire pouvant par la suite provoquer un effet Allee démographique. Ces mécanismes agissent sur la survie et la reproduction des individus et font varier la valeur sélective d’une population en fonction de sa taille. Pour autant cet effet n'affecte pas l'ensemble du règne du vivant, ce n'est pas un phénomène universel, loin de là. De plus il est très difficile de le mettre en évidence car on ne l'observe que dans les populations à faible densité.
Aspect génétique
Dans une population à faible densité la consanguinité risque d'être plus importante, ce qui affecte la mortalité et la reproduction des individus, aboutissant à une diminution du taux de croissance dans ces populations (inbreeding depression).
Aspect comportemental
Un effet Allee peut aussi être causé par des comportements coopératifs. Chez les espèces sexuées notamment il est nécessaire à chaque individu de trouver un partenaire pour la reproduction. Plus une population est petite plus il est difficile d'entrer en contact avec un partenaire potentiel. Cela a pour conséquence de diminuer le taux de reproduction dans ces populations, à faible densité. Ce phénomène est appelé « mate limitation » (limitation par le partenaire). On l'observe en particulier chez les invertébrés aquatiques, car la reproduction passive dans l'eau rend difficile la rencontre des gamètes quand les individus sont peu nombreux. Il en est de même pour les plantes terrestres pour lesquelles on parle de limitation de pollen[3].
Dans les populations vivant en groupes sociaux, les comportements comme l’élevage des jeunes, la recherche de nourriture ou encore la défense en groupe, sont plus efficaces au sein d’un grand groupe d’individus. Les Lycaons, par exemple, vivent en groupes sociaux ; plus ces groupes sont importants mieux ils se protègent, car certains adultes se consacrent à l'élevage et à la protection des jeunes. De plus, de grands groupes chassent plus efficacement. Cela a un effet positif sur la survie et donc sur le taux de croissance de la population[2].
D’autres mécanismes n’impliquent pas de comportements coopératifs. La seule présence de nombreux individus améliore la survie de ces individus. Par exemple au sein d’un large groupe le risque de prédation pour chaque individu est plus petit que dans un groupe réduit[3]. Le phénomène de saturation du prédateur requiert aussi un grand nombre d'individus.
Aspect physiologique
Il a été mis en évidence que la présence de nombreux individus en un lieu peut altérer cet environnement (biotique ou abiotique) et cela de manière à favoriser les conditions de vie de ces individus. On parle aussi de construction de niche, observée par exemple chez des plantes faisant de l’allélopathie ou qui acidifient leur milieu (Tsuga heterophylla p.ex.).
Aspect anthropogénique
Certains chercheurs dont, en France, Franck Courchamp, estiment que l'exploitation par l'homme peut non seulement révéler un effet Allee mais aussi en être la cause. On parle alors d'effet « rareté »[5]. En effet, comme le sous-entend le proverbe « ce qui est rare est cher » une valeur arbitraire élevée peut être attribuée par l'homme à des espèces vivantes rares ou à des éléments de leur environnement.
Dès lors un effet Allee anthropogénique peut se manifester dans les populations soumises à des fortes pressions anthropiques par diverses activités, comme :
- collections d'animaux, de plantes rares (cactées, orchidées, insectes...)
- phénomène NAC (Nouveaux Animaux de Compagnie) : animaux exotiques recherchés par certains particuliers)
- chasse d'animaux rares et recherche de trophées
- médecine traditionnelle (corne de rhinocéros...)
- matières et bois précieux (exploitation du bois de rose à Madagascar dans les réserves naturelles, ivoire...)
- écotourisme
- marchés du luxe (caviar...)
Cet effet pose des problèmes particuliers en conservation de la nature, laissant penser que le fait de déclarer une espèce « rare » pourrait la mettre en danger.
Conséquences
Évolution
Dans une population où il y a un effet Allee, la valeur sélective individuelle augmente avec la densité. En conséquence les caractères facilitant l'agrégation devraient être favorisés par sélection naturelle lors de l'évolution.
Exploitation et conservation
L'exploitation rationalisée des populations naturelles (ex : la pêche intensive) est généralement conduite de manière à maintenir ces populations à la densité de K/2, densité pour laquelle le taux de croissance de la population est maximal. Or, si la population exploitée manifeste un effet Allee, à K/2 le taux de croissance n'est plus maximal, mais beaucoup plus faible et la population ne se régénère pas assez vite pour compenser l'exploitation. C’est ce qui s’est passé avec certains stocks de poissons exploités au MSY (« maximum sustainable yield » ou rendement maximal soutenable, déterminé à partir de K/2). Il faut donc prendre en compte l’effet Allee lors de la gestion des populations naturelles[6].
L’effet Allee peut également diminuer le risque d’invasions biologiques. En effet si une espèce exotique arrive dans un nouveau milieu en effectif trop faible, en dessous du seuil critique (A) prédit par le modèle de l’effet Allee, elle ne pourra pas s’établir. Ceci pourrait expliquer pourquoi uniquement une petite fraction des espèces introduites réussit à se maintenir et/ou à envahir un nouveau milieu.
Enfin, une petite population d’une espèce qui montre un fort effet Allee a une probabilité élevée de s’éteindre. Ainsi on peut définir, pour certaines espèces rares, une taille de population minimale MVP (« minimum viable population »), qui correspond à la densité critique nécessaire à sa persistance dans le milieu, pour une aire de répartition donnée, qui permet de tenir compte de l’effet Allee dans les modèles de dynamique de ces populations[2].
Voir aussi
Articles connexes
- Biologie de la conservation
- Comportement animal
- Démécologie
- Dynamique des populations
- Espèce menacée
- Extinction des espèces
- Liste rouge de l'UICN, sur les espèces animales et végétales menacées d'extinction
- Plan de restauration ; gestion restauratoire
- Psychologie sociale
- Réintroduction
- Statut de conservation
Liens externes
- (en) Rapport de synthèse Biodiversité de l'Evaluation des écosystèmes pour le Millénaire (EM, 2005) traitant de l'extinction des espèces.
- (fr) Résumé en français de ce Rapport de synthèse du EM par GreenFacts.
- Notes de cours de paléontologie de Biodeug.
- (fr) [PDF] Crises biologiques et extinctions de masse [(fr) lire en ligne], sur le site Geopolis, confédération française des acteurs des sciences de la terre.
- (en) The Nature education : Knowledge Project : Allee effects Article de synthèse très complet sur le sujet.
Bibliographie
- Richard Leakey, La Sixième Extinction — Évolution et catastrophes [« The Sixth Extinction, Patterns of Life and the Future of Humankind »], Flammarion, (ISBN 2-08-081426-5)
- Philippe Dubois, Vers l'ultime extinction? : la biodiversité en danger, La Martinière, , 191 p. (ISBN 978-2-7324-3090-4)
- Blackwood J.C, Berec L, Yamanaka T, Epanchin-Niell R.S, Hastings A & Liebhold A.M (2012) Bioeconomic synergy between tactics for insect eradication in the presence of Allee effects. Proceedings of the Royal Society of London B: Biological Sciences, 279(1739), 2807-2815.
Notes et références
- University of Chicago Library (2006) Guide to the Warder Clyde Allee Papers 1894-1980.
- Courchamp F., Cutton-Brock T., Grenfell B. (1999) Inverse density dependance and the Allee effect, TREE vol 14, no. 10.
- Kramer A.M., Dennis B., Liebhold A.M., Drake J.M. (2009) The evidence for Allee effects, Popul Ecol 51, 341-354.
- Flower M.S., Ruxton G.D. (2002) Population Dynamic Consequences of Allee Effects, J. theor Biol. 215, 39-46.
- Franck Courchamp, "Un dilemme pour la protection des espèces", article dans La Recherche.
- Stephens P.A., Sutherland W.J. (1999) Consequences of the Allee effect for behaviour, ecology and conservation, TREE vol 14 no. 10, 401-405.
{kind=link}