Lordose (comportement sexuel)
Dans le comportement sexuel des mammifères, la lordose correspond chez les femelles à une position sexuelle réflexe qui permet la copulation.
Au niveau pathologique, l'hyperlordose correspond à une courbure excessive de la colonne vertébrale.
La lordose est une courbe concave décrite par la colonne vertébrale dans le plan sagittal. Elle s'oppose à la cyphose, où la colonne décrit une courbe convexe. Chez l'homme la colonne vertébrale a quatre courbures naturelles : la lordose cervicale (aspect creusé du cou), suivie de la cyphose thoracique (la bosse du dos), puis la lordose lombaire (creux en bas du dos), enfin la cyphose sacrale/coccygienne. Une mauvaise posture se traduit par des modifications des courbes décrites par la colonne : des vertèbres en cyphose peuvent décrire une lordose et le terme désigne alors un état pathologique (on parle alors de lordose excessive ou hyperlordose).
La lordose désigne également chez les mammifères femelles un réflexe inné de courbure de la colonne vertébrale pendant la copulation, pour présenter le vagin au mâle et ainsi faciliter la pénétration. Ce réflexe copulatoire est précablé dans le cerveau, contrôlé par les hormones dans l'hypothalamus[1], déclenché par des contacts physiques sur les flancs, la croupe ou le périnée[2], et facilité par les stimulations vaginales[3] et par les phéromones sexuelles[4].
Réflexe sexuel des mammifères femelles
 La lordose est particulièrement prononcée chez les félidés
La lordose est particulièrement prononcée chez les félidés.jpg.webp) La lordose est une position indispensable à la copulation
La lordose est une position indispensable à la copulation La lordose est innée chez tous les mammifères non-primates
La lordose est innée chez tous les mammifères non-primates Lordose chez les éléphantidés
Lordose chez les éléphantidés
Chez les mammifères femelles, la lordose est un réflexe sexuel moteur, complexe et crucial.
On observe qu’il est fonctionnel et indispensable chez tous les mammifères non-primates (rongeurs, canidés, félidés, bovidés, équidés…), mais qu’à partir des primates et surtout des hominidés (chimpanzés, orangs-outans, gorilles, humains), les réflexes innés deviennent secondaires. Chez l’être humain, le réflexe n'est pas fonctionnel[5].
Intégration du réflexe de lordose dans le comportement de reproduction
Les études scientifiques récentes montrent que l'organisation neuroanatomique générale des mammifères est spécifiquement conçue pour la copulation hétérosexuelle[6]. En simplifiant, il existe trois grands circuits neurobiologiques : 1) les circuits olfactifs (flèches rouges, schéma ci-dessous), à l'origine de l'excitation sexuelle et de l'orientation sexuelle ; 2) les circuits des réflexes sexuels (lordose, érection, éjaculation… flèches orange), qui permettent la copulation ; et 3) les circuits des récompenses sexuelles (système de récompense associé au pénis/clitoris – flèches bleues), qui sont impliqués dans les apprentissages sexuels (en particulier de la motivation sexuelle)[5]. L’analyse des circuits sexuels montre que le réflexe de lordose correspond, chez la femelle, à la partie motrice du comportement de reproduction.
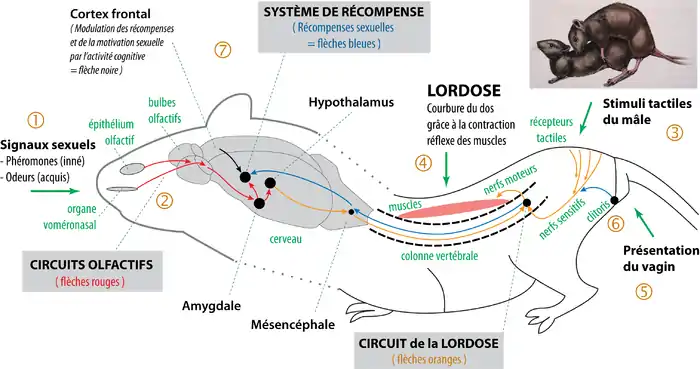
En simplifiant, on peut schématiser les principales phases du comportement de reproduction. 1) Les odeurs et surtout les phéromones permettent d'échanger des signaux sexuels entre les éventuels partenaires[7]. 2) Les circuits olfactifs (flèches rouges) permettent en particulier de reconnaître le partenaire du sexe opposé[8] - [9] - [10] et de déclencher l'excitation sexuelle[11] - [12], ce qui induit la lubrification vaginale, l'érection et la copulation. 3) Lorsque le mâle monte la femelle, les stimuli tactiles du mâle sur la croupe de la femelle déclenchent le réflexe de lordose[13] - [14]. 4) Les circuits de la lordose (flèches orange) provoquent la courbure du dos, ce qui permet de bien présenter le vagin au mâle[13] - [14]. 5) La présentation du vagin facilite le contact tactile du pénis avec la région génitale, ce qui déclenche les mouvements réflexes du bassin du mâle (poussées pelviennes). Après l'intromission, les mouvements du pénis dans le vagin déclenchent le réflexe d'éjaculation[15] - [16]. 6) Les stimulations tactiles du clitoris (et du pénis pour le mâle) durant la copulation remontent jusqu'au cerveau (flèches bleues)[17]. 7) L'activation du système de récompense induit des apprentissages qui optimisent la copulation, en particulier le développement de la motivation sexuelle[18]. De plus, des signaux olfactifs, auditifs et visuels perçus au cours de la copulation peuvent devenir par conditionnement des signaux sexuels[19], ce qui optimise les signaux phéromonaux innés[11]. Il existe ainsi, dans l'organisation neurobiologique innée de l'organisme, un véritable comportement de reproduction hétérosexuel chez les mammifères non-primates[6] - [5].
Organisation neurobiologique du réflexe de lordose
L'analyse détaillée de l'organisation neurobiologique du réflexe moteur complexe de la lordose montre que ce réflexe est spécifiquement précablé pour présenter le vagin au mâle lors de la copulation, et uniquement quand la femelle est fécondable. (cf figure ci-dessous)

Le réflexe moteur de la lordose est principalement précablé dans la moelle épinière, au niveau des vertèbres lombaires L1, L2, L5 et L6 et des vertèbres sacrées S1[14]. Dans le cerveau, plusieurs régions modulent le réflexe de lordose. Les noyaux vestibulaires et le cervelet, via le faisceau vestibulaire, envoient des informations qui permettent de coordonner le réflexe de lordose avec l'équilibre postural. Et surtout, l'hypothalamus ventromédian envoie des projections qui inhibent le réflexe au niveau médullaire[13]. Pour cette raison, en général, le réflexe de lordose n'est pas fonctionnel. Les hormones sexuelles contrôlent la reproduction et coordonnent les activités sexuelles avec l'état physiologique. En schématisant, à la période de reproduction, et quand un ovule est disponible, les hormones (en particulier les œstrogènes) induisent simultanément l'ovulation et l'œstrus (les « chaleurs »). Sous l'action des œstrogènes dans l'hypothalamus, le réflexe de lordose n'est plus inhibé[1]. La femelle est prête pour la copulation et la fécondation. Au cours de la copulation, quand un mâle s'approche de la femelle, les phéromones du mâle (partie 1 du schéma ci-dessus), sont détectées par les circuits olfactifs (partie 2). Les signaux phéromonaux stimulent, entre autres, l'hypothalamus, ce qui facilite le réflexe de lordose[4]. Puis, quand le mâle monte la femelle (partie 3), les stimuli tactiles sur les flancs, la croupe et le périnée de la femelle sont transmis via les nerfs sensitifs dans la moelle épinière. Dans la moelle, ils sont intégrés avec les informations provenant du cerveau, puis, en général, un influx nerveux est transmis dans les muscles via les nerfs moteurs. La contraction des muscles longissimus et transverso-spinalis provoque la courbure de la colonne vertébrale (partie 4)[14]. La position de lordose qui en résulte permet de bien présenter le vagin au mâle (partie 5), ce qui facilite l'intromission du pénis. Puis, au cours de l'intromission, les sensations tactiles et profondes provenant de la région génitale et du clitoris accentuent le réflexe de lordose (partie 6) [3]. On observe ainsi que l'organisation physiologique et neurobiologique du réflexe de lordose est adaptée à la copulation hétérosexuelle.
Évolution de la lordose
Des rongeurs à l'être humain, le développement majeur du cerveau entraîne plusieurs modifications du contrôle de la sexualité, dont celui de la lordose[20].
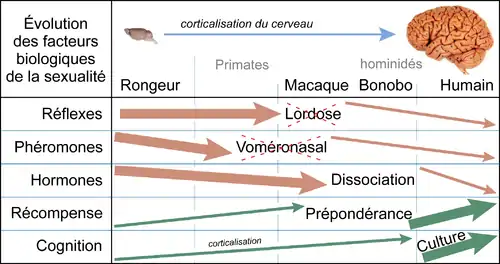
Ces modifications neurobiologiques induisent une évolution du comportement sexuel stéréotypé des mammifères non-primates vers « l'étonnante variété des comportements sexuels humains »[6]. C’est à partir des anthropoïdes (les simiens, ou les singes) qu’on observe une absence progressive de la lordose réflexe, parallèlement à l’absence progressive d’effets hormonaux importants et d’œstrus[21]. Chez les cercopithèques (macaques...), le réflexe de lordose est encore fonctionnel, mais il est intégré dans des activités sexuelles et aussi sociosexuelles, comme la présentation. Cette activité sociosexuelle de présentation (qui correspond aux mêmes séquences que la lordose : courbure du dos et déplacement latéral de la queue, ce qui présente la région génitale), ne dépend plus d’un contrôle hormonal, mais est réalisée en fonction de la situation sociale[22]. Chez les hominidés, la lordose et l’immobilisation sont absentes, même durant la phase péri-ovulatoire. La réceptivité de la femelle n’est ni obligatoire, ni passive. Elle peut activement éviter ou refuser le mâle, ou terminer la monte[21].
Dans l'espèce humaine et chez les hominidés
En raison de ces évolutions biologiques, le réflexe copulatoire de lordose n'est plus fonctionnel chez l'être humain et les autres hominidés[20]. Chez les humains, le réflexe moteur de la copulation a été remplacé par de nouvelles activités sexuelles, dont certaines n'ont plus aucun rapport avec la copulation et la fécondation. Contrairement à la lordose, les activités sexuelles humaines (baiser, masturbation mutuelle, fellation, coït en face-à-face…) ne sont plus des réflexes moteurs, mais principalement des activités volontaires et apprises, réalisées pour obtenir des récompenses sexuelles[23] - [24] - [6].
Références
- (en) Flanagan-Cato L.M. Sex differences in the neural circuit that mediates female sexual receptivity. Frontiers in Neuroendocrinology, 32(2):124-136, 2011.
- Pfaff D.W., Schwartz-Giblin S., MacCarthy M.M., Kow L.M. Cellular and molecular mechanisms of female reproductive behaviors. in Knobil E., Neill J.D. (Eds). The physiology of reproduction. Raven Press, 2nd edition, 1994
- Gonzalez-Flores O., Beyer C., Lima-Hernandez F.J., Gomora-Arrati P., Gomez-Camarillo M.A., Hoffman K., Etgen A.M. Facilitation of estrous behavior by vaginal cervical stimulation in female rats involves alpha1-adrenergic receptor activation of the nitric oxide pathway. Behavioural Brain Research, 176(2):237-243, 2007
- (en) Haga S., Hattori T., Sato T., Sato K., Matsuda S., Kobayakawa R., Sakano H., Yoshihara Y., Kikusui T., Touhara K. The male mouse pheromone ESP1 enhances female sexual receptive behaviour through a specific vomeronasal receptor. Nature, 466(7302):118-122, 2010.
- (fr) Wunsch S. Comprendre les origines de la sexualité humaine. Neurosciences, éthologie, anthropologie. [PDF] L'Esprit du Temps, 2014.
- (en) Agmo A. Functional and dysfunctional sexual behavior
- (en) Keller M., Bakker J. Pheromonal communication in higher vertebrates and its implication on reproductive function. Editorial. Behavioural Brain Research, 200(2):237-238, 2009.
- (en) Stowers L., Holy T. E., Meister M., Dulac C., Koentges G. Loss of sex discrimination and male-male aggression in mice deficient for TRP2, Science, 295(5559):1493-1500, 2002.
- Agustin-Pavon C., Martinez-Garcia F., Lanuza E. Focal lesions within the ventral striato-pallidum abolish attraction for male chemosignals in female mice. Behavioural Brain Research, 259:292-296, 2014
- (en) Dulac C., Torello A. T. Molecular detection of pheromone signals in mammals: from genes to behaviour, Nat. Rev. Neurosci., 4(7):551-562, 2003.
- (en) Moncho-Bogani J., Lanuza E., Hernandez A., Novejarque A., Martinez-Garcia F. Attractive properties of sexual pheromones in mice: innate or learned? Physiology & Behavior, 77(1):167-176, 2002.
- (en) Yoon H., Enquist L. W., Dulac C. Olfactory inputs to hypothalamic neurons controlling reproduction and fertility, Cell, 123(4):669-682, 2005.
- (en) Kow L.M., Florea C., Schwanzel-Fukuda M., Devidze N., Kami K.H., Lee A., Zhou J., Maclaughlin D., Donahoe P., Pfaff D. Development of a sexually differentiated behavior [lordosis] and its underlying CNS arousal functions. Curr. Top. Dev. Biol., 79:37-59, 2007
- (en) Pfaff Donald W., Schwartz-Giblin Susan, MacCarthy Margareth M., Kow Lee-Ming. Cellular and molecular mechanisms of female reproductive behaviors, in Knobil Ernest, Neill Jimmy D. The physiology of reproduction, Raven Press, 2nd edition, 1994
- (en) Allard J., Truitt W. A., McKenna K. E., Coolen L. M. Spinal cord control of ejaculation, World J. Urol., 23(2):119-126, 2005.
- (en) Coolen L. M. Neural control of ejaculation, J. Comp Neurol., 493(1):39-45, 2005.
- (en) Matsumoto J., Urakawa S., Hori E., de Araujo M.F., Sakuma Y., Ono T., Nishijo H. Neuronal responses in the nucleus accumbens shell during sexual behavior in male rats. The Journal of Neuroscience, 32(5):1672-1686, 2012.
- (en) Cibrian-Llanderal T., Tecamachaltzi-Silvaran M., Triana-Del R.R., Pfaus J.G., Manzo J., Coria-Avila G.A. Clitoral stimulation modulates appetitive sexual behavior and facilitates reproduction in rats. Physiology & Behavior, 100(2):148-153, 2010.
- (en) Pfaus J.G., Kippin T.E., Coria-Avila G.A., Gelez H., Afonso V.M., Ismail N., Parada M. Who, what, where, when (and maybe even why)? How the experience of sexual reward connects sexual desire, preference, and performance. Archives of Sexual Behavior, 41(1):31-62, 2012.
- Wunsch S. Phylogénèse de la sexualité des mammifères. Analyse de l'évolution des facteurs proximaux. Sexologies, 26(1):3-13, 2017.
- Dixson A.F. Primate sexuality: Comparative studies of the Prosimians, Monkeys, Apes, and Human Beings. Oxford University Press, 2nd edition, 2012
- Hanby J.P. Sociosexual development in primates. in Bateson P.P. (Ed). Perspectives in Ethology, volume 2. Plenum Press, (1):1-67, 1976
- (fr) Wunsch S. Thèse de doctorat sur le comportement sexuel [PDF] EPHE-Sorbonne, Paris, 2007.
- Georgiadis J.R., Kringelbach M.L., Pfaus J.G. (2012) Sex for fun: a synthesis of human and animal neurobiology. Nat. Rev. Urol., 9(9):486-498.