Gènes KNOX
Les gènes KNOX (knotted-like homeobox) sont connus pour avoir un rôle dans le développement des plantes. Ce sont des gènes homéotiques dont les produits ont pour fonction l’élaboration et la conservation des structures aériennes chez toutes les plantes terrestres et certains groupes d’algues vertes. Les gènes et leur produits (des protéines) sont de type TALE et sont subdivisés en classes, soient les gènes KNOX de classe I, de classe II et les KNATM[1]. La classification se base sur la séquence, les domaines (modules protéiques retrouvés dans plusieurs protéines et dont la fonction est constante), la disposition des introns et exons[2] - [3] entre autres.
Les produits de gènes homéotiques, dont les gènes KNOX, sont des facteurs de transcription dont la fonction est la régulation (activation/répression) des gènes de développement. Ils contiennent généralement tous un homéodomaine, domaine spécifique de liaison à l’ADN dont la séquence est bien conservée chez les Animaux, les Plantes et les Mycètes (ie. chez les Eucaryotes).
L’équivalent des gènes homéotiques chez les animaux sont les gènes Hox (HOX genes).
Les études dans le règne végétal sont principalement effectuées chez l’organisme modèle Arabidospis thaliana. Le séquençage du génome a révélé, chez cette espèce, 110 gènes homéotiques, mais les experts mentionnent que ce nombre devrait être assez constant chez toutes les plantes[3].
Structure et fonctions
L’homéoboîte (normalement présente dans les gènes homéotiques) code typiquement une séquence de 180 paires de nucléotides dont l’homéodomaine protéique résultant est constitué de 60 acides aminés ayant pour fonction la liaison à l’ADN[2]. La liaison a pour fonction la régulation (activation ou inhibition) de gènes en cascade, dont l’expression résulte en la formation d’une structure entière et fonctionnelle (e.g. un bras fonctionnel chez les animaux, ou une tige fonctionnelle chez les plantes terrestres).
La liaison à l’ADN de l’homéoprotéine seule ne procure pas assez de spécificité pour réguler l’activité des gènes récepteurs. Ainsi, il y a nécessité de l’ajout de domaines et de l’interaction complexe avec des cofacteurs dont les sites peuvent être a faible ou forte affinité[3] (donc où la liaison des protéines peut respectivement se faire avec un plus grande variété de séquence de l’ADN ou une plus faible variété de séquences de l’ADN, voire une seule séquence), en plus de l’arrangement de l’ADN elle-même qui, tous ensemble, procurent une grande spécificité protéique.
Un phénomène couramment observé dans l’évolution est tendance à la multiplication du nombre de gènes homéotiques, donc de protéines homéotiques, avec la complexification et la multiplication fonctionnelle de la vie[1] (des eucaryotes unicellulaires aux plantes et animaux multicellulaires modernes). La section « Classification » traitera plus amplement de l’origine et des mécanismes de la diversification des protéines homéotiques, particulièrement des protéines KNOX.
Par ailleurs, il existe une super-classe d’homéoprotéines nommée TALE (three amino-acid loop extension, pouvant être traduit par « extension en boucle de trois acides aminés »). Cette classe code donc pour des protéines contenant un homéodomaine de 63 acides aminés, en opposition à l’homéodomaine "typique" de 60 acides aminés. Les protéines TALE sont sub-divisées en deux familles : BELL et KNOX.
Dans la famille KNOX, la majorité des protéines contiennent à la fois un domaine KNOX1, KNOX2, ELK et l’homéoboîte. On distingue toutefois les protéines ne contenant que le domaine KNOX1 et KNOX2, qu’on nomme KNATM[1]. Voir le tableau « Domaines et fonction des protéines KNOX ».
| Domaine | Fonction | Apparition | Retrouvé dans les protéines KNOX |
|---|---|---|---|
| KNOX 1/A | peu d’études existent - peuvent avoir un lien avec les homo et hétéro-dimérisations (BELL) procurant la spécificité de liaison à l’ADN, ou agir comme compétiteur au site de liaison à l’ADN[2] | avant la divergence des animaux et des plantes (sous la forme ancestrale MEINOX), modifié en KNOX chez tous les végétaux[2] | Classe I, Classe II et KNATM |
| KNOX 2/B | Classe I, Classe II (dans cette classe, insertion de 30 acides aminés[2] et KNATM | ||
| KNOX C | chez les plantes à fleur, avant la divergence des eudicotylédones et monocotylédones[2] | Classe II seulement | |
| ELK | localisation nucléaire pour la protéine [4] | avant la divergence des animaux et des plantes | Classe I et Classe II seulement |
| Homéoboîte de type TALE (présentant une insertion homologue de trois acides-aminés) | liaison spécifique à l’ADN[1] | chez les eucaryotes, avant la divergence des animaux, mycètes et plantes | Classe I et Classe II seulement |
KNOX et le développement
Les plantes terrestres présentent à ce jour entre 5 et 13 protéines KNOX dans leur génome[2]. Parmi ces protéines, on dénote trois groupes : la classe I, la classe II et les KNATM (ces dernières ne sont pas des protéines homéotiques à proprement parler car elles ne présentent pas d’homéodomaine).
Classe I
Les gènes des protéines de la classe I, chez les plantes à fleur dont l’espèce modèle Arabidospis thaliana, sont sub-divisés en différents groupes : KNAT2/6, STM (shoot meristemless) et KNAT 1[2] - [1]. Les protéines de la classe I sont exprimées dans la région restreinte du méristème apical caulinaire (MAC, ou SAM en anglais). Ils régulent la formation, la maintenance, le bon fonctionnement du MAC, chez toutes les plantes vasculaires, incluant les Lycophytes, et Euphylophytes (fougères), Angiospermes et Gymnospermes[5]. La classe I jouerait un rôle dans le maintien de la bonne identité cellulaire dans le tissu méristématique durant la vie de la plante et aurait également un lien avec le développement des fleurs et des feuilles.
Par exemple, on note que le premier gène homéotique à être exprimé dans les plantes est le gène Kn1 (chez Arabidospis thaliana, on utilise l’appellation KNAT 1), au niveau de la germination de la graine. On le retrouve ensuite normalement dans le site du MAC. On constate que dès qu’il y a initiation de la feuille en position apicale, le gène est réprimé[6], mais que dans la formation de structure en position latérale, le gène est exprimé[7]. Puis, la mutation des gènes STM résulte en l’absence complète du MAC chez A. thaliana[8], indiquant qu’ils ont un rôle crucial dans la formation de cette structure.
Le méristème apical caulinaire
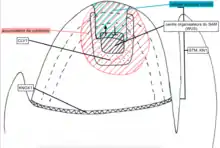
Le méristème apical caulinaire ou MAC est la structure initiatrice du développement de toutes les structures caulinaires et des organes chez les plantes terrestres (ou plantes vasculaires). Très dynamique, il est formé de 1 à 3 cellules indifférenciées et multipotentes (des cellules souches), dont l’élongation et la différenciation résultent en la tige, les feuilles et fleurs ainsi que d’autres structures reproductrices de la plante.
Bien que chez les différents groupes de plantes, le MAC présente différentes zones et différentes stratifications, les experts s’entendent pour dire qu’il possède toujours la même fonction. On pense toutefois que son apparition commune chez les différents groupes est due à l’évolution parallèle (trois différentes apparitions, respectivement chez les fougères, les Lycophytes et les plantes à graines) et ne serait donc pas apparu avant la divergence des différents groupes[5].
La structure MAC comprend un centre organisateur, une région d’accumulation de cytokinine (phytohormone), une zone centrale où le gènes CLV3 (famille CLAVATA) est exprimé (division cellulaire de cellules souches), des régions d’expression de différents gène de la classe TALE.
La régulation l’activité du MAC est assurée par les protéines TALE, voir plus bas dans la section « Régulation ».
Les experts mentionnent que la densité des plasmodesmes sur les parois cellulaires des cellules du MAC varie entre les différents groupes de végétaux. La densité (nombre par unité d’aire) de plasmodesmes peut en effet être qualifiée de haute ou basse, et cette variable joue un rôle important dans la régulation des activités, notamment dans le transfert entre cellules d’ARNm, de facteurs de transcription, phytohormones et morphogènes[5].
Classe II
Les gènes des protéines de la classe 2, chez les plantes à fleur, sont par ailleurs subdivisés en deux groupes : KNAT7 et KNAT3/4/5[2]. La classe II des protéines KNOX est mieux conservée chez les plantes terrestre, ce qui indique qu’elle est soumise à une plus grande force sélective. Elle est connue pour être exprimée de manière plus étendue dans la plante, c’est-à-dire incluant les racines et les feuilles[8], ainsi que dans des tissus haploïde, c’est-à-dire reproducteurs.
Peu d’études ont été menées pour élucider leurs rôles. Des expériences de knock-out (où le gène cible est éliminé du génome pour pouvoir en déterminer les effets résultants sur l’organisme) ont été effectuées et les plantes dont on élimine l’expression des protéines KNOX de classe II ne montrent pas un phénotype anormal[8]. Ainsi, leurs rôles n’ont pu être clairement démontrés. Malgré tout, des études ont démontré que ces protéines semblent favoriser la transition du tissu somatique vers le tissu reproducteur[1], semblent avoir une fonction dans la régulation de la déposition de la paroi secondaire des cellules végétales, dans l’établissement du site d’initiation de la feuille et semblent également avoir un lien avec la régulation de la réponse à l’acide abscissique (une phytohormone) dans les processus initiaux de développement de la graine[1].
KNATM ou "Classe III"
Les KNATM sont impliquées dans la régulation de la transcription, la détermination de l’axe de la feuille, l’initiation du méristème floral, dans les hydathodes (pores sur l’écorce sécrétant des liquides de composition variable) et on note que son expression est strictement exclue du MAC. Ses domaines KNOX 1 et 2 lui permettent, comme dans le cas des KNOX de classe I et II, d’interagir avec les protéines BELL. Ainsi, on présume que son importance réside dans le rôle de cofacteur de transcription ou comme régulateur de l’expression des autres protéines de la famille TALE. Chez Glycine max, une eudicotylédone, on dénote la présence de nombreux paralogues (copies du/des gènes, dans une espèce, issues de la duplication antérieure de leur séquence) des protéines KNATM, résultant d’événement(s) de duplication. Chez cette espèce, les experts constatent que ces paralogues de KNATM sont très modestement exprimées dans la plante. En effet, leur expression se résumerait seulement à l’axe de la plumule, au moment de la germination[1] - [9].
Classification et ancestralité
On oppose la super-classe TALE et « non-TALE ». Chez les plantes, la super-classe TALE contient les classes BELL et KNOX et la superclasse « non-TALE » compte 12 familles, soient : PLINC, PHD, WOX, NDX, SAWADEE, PINTOX, HD-ZIP I,II, III et IV, LD, DDT.
Cette classification[3] se base sur les domaines constituant les protéines et la séquence de l’homéodomaine dans l’ADN[2]. La super-classe de protéines TALE est retrouvée chez les plantes comme chez les animaux et les eucaryotes primitifs, ce qui révèle une mutation dans l’homéoboîte survenue avant la divergence (séparation) de ces trois groupes[3]. Les protéines KNOX et BELL sont semblables dans leurs fonctions et leur structure, si bien qu’elles s’associent souvent sous forme d’"hétérodimères". Certaines protéines KNOX montrent également la capacité de former des homodimères, lorsque deux protéines du même type s’associent ensemble[9]. Voir la section "Régulation" pour plus de détails. Ces deux groupes sont apparus avant même la divergence des différents groupes de plantes. Leur ressemblance est probablement due à une duplication du domaine MEINOX ("ancêtre" de BELL et KNOX), puis à l’accumulation de mutations ayant mené aux BELL et KNOX[2]. Leur séquence, pourtant toujours semblable, permet l’association en dimères.
La famille des protéines KNOX est monophylétique de même que la Classe I et II, comme le démontrent des analyses phylogénétiques[3] - [1] - [2]. Ces dernières classes sont retrouvées chez toutes les plantes terrestres. Les gènes KNOX des algues vertes (Chlorophyta, Charophyta) toutefois ne peuvent être associés à l’une ou l’autre de ces deux classes, ils forment donc leur clade propre. Par conséquent, les experts considèrent que les protéines KNOX sont apparues chez les algues vertes et qu’elles ont dupliqué en classes I et II telles que nous les connaissons, après la divergence des plantes terrestres, il y a environ 400 millions d’années. Malgré le fait que les protéines KNOX des plantes vertes ancestrales forment leur propre groupe, elles ont plus de ressemblance avec la classe I des plantes terrestres. Ainsi, la classe I serait probablement la forme ancestrale des gènes KNOX des plantes terrestres et autrefois exprimée partout dans la plante, incluant les racines et les inflorescences[6]. La duplication permit l’accumulation de mutations sur les copies résultantes, jusqu’à se rapprocher des séquences protéiques des classe I et II modernes. Dans les différentes espèces, puisque les protéines de la classe I nécessitent plus de spécificité de régulation, leur séquence varie plus. Puis, on mentionne que l’expression de la classe II est plus étendue donc ne nécessite pas autant de spécificité et leur séquence est restée plus ou moins constante dans les différentes plantes terrestres[2].
Des études ont émis l’idée que les gènes KNOX de classe I étaient ancestralement exprimés dans toute la plante, de manière non spécifique. Puis, la spécialisation de la fonction de la classe II aurait permis l’inhibition d’expression de la classe I à l’extérieur du MAC dans les plantes[1] - [6] - [10]. Toutefois les recherches mentionnent que le manque de données rend difficile l’analyse phylogénétique des gènes KNOX chez les plantes, et ne permet pas une compréhension complète de l’origine, soit paralogue soit orthologue des gènes.
Comme mentionné ci-haut dans la section « Structure et fonctions », l’augmentation de la diversité de protéines, dans la famille KNOX comme dans d’autres, permet souvent aux organismes d’également gagner une plus grande diversité de fonctions. Une étude mentionne qu’il y eut deux événements d’expansion de la famille KNOX (par duplication et accumulation de mutations dans les copies paralogues)[3]. Dans un premier temps, elle se produit dans les mousses (Bryophyta), au moment de la transition vers la vie terrestre. Cette expansion fut avantageuse car elle permit l’accumulation de mutations qui ont permis de répondre aux nombreux défis qu’impose la vie sur la terre ferme[1] (expositions aux UV, risque de dessiccation, lutte contre la gravité, stratégies de reproduction et autres).
Une deuxième expansion se produit au moment de la transition des Lycophytes vers la branche qui mena aux Angiospermes. Elles ont probablement contribué à la complexification des feuilles et autres tissus. D’autres exemples de complexification et multiplication protéique existent chez les Angiospermes, par exemple dans le cas des gènes homéotiques MADS qui ont permis, pour leur part, la diversification des stratégies de reproduction (développement des verticiles floraux)[1].
Les KNATM ne sont retrouvées que chez les Eudicotyldones[1] - [9], ce qui suggère qu’ils proviennent de la duplication suivie de la perte des autres domaines typiques des protéines KNOX.
Régulation
Les protéines KNOX peuvent interagir avec les BELL, notamment en hétérodimérisant, c’est-à-dire en s’associant de manière très spécifique. Par exemple, dans le cas des protéines BEL1 et STM, leur région d’expression dans le MAC est en partie superposée, ce qui laisse croire qu’ils hétérodimérisent pour réguler la transcription dans cette région.
Les protéines CLAVATA (CLV), dont l’expression des CLV1 et CLV3 est montrée dans le du MAC (section MAC), semblent réguler l’accumulation de cellules souches dans le MAC, et entraîner leur différenciation. Selon toute évidence, elles jouent un rôle antagoniste à celui du STM, qui pour sa part semble entraîner la prolifération des cellules souches dans le MAC et ralentir la différenciation[8] - [9].
Il a été proposé que, dans l’événement d’initiation de la feuille, au niveau du primordia, les gènes KNOX soient régulés par les protéines PHANTASTICA et MGOUN. Respectivement, il semble qu’elles ont un rôle dans la régulation des KNOX pour la détermination de l’axe adaxial-abaxial et le nombre et la grosseur du/des méristème(s)[8].
Dans une feuille en croissance ou mature, ce sont plutôt les gènes BOP1 et BOP2 qui répriment l’expression des KNOX de classe I.
Les protéines homéotiques, dont les KNOX et BELL, jouent deux rôles particulièrement importants chez les plantes. Le premier est un lien étroit avec la régulation hormonale. Par exemple, les KNOX assurent la régulation hormonale des cytokinines (promotion de la division cellulaire) et gibbérellines (élongation et initiation du primordia foliaire), des phytohormones, dans le signalement du MAC. En effet, il fut démontré que la cytokinine et la gibbérelline sont régulées par des gènes KNOX de classe I[11], qui en empêchent l’expression dans le MAC. Un autre exemple est celui de l’hormone auxine qui inhibe l’expression des KNOX dans la zone de formation des organes (hors du MAC). Puis, l’éthylène pourrait également avoir un rôle de régulation dans le MAC, car on observe des interactions avec la protéine KNOX2[9].
Le deuxième rôle est qu’elles interagissent avec les protéines MADS (acronyme pour MCM1, AGAMOUS, DEFICIENS, SRF). Dans le cas des KNOX, on n’a pas fermement compris les interactions avec les gènes MADS, mais il est fortement probable qu’ils s’influencent mutuellement dans la cascade d’événements qui précède le développement des fleurs[11].
Notes et références
- Gao, J., Yang, X., Zhao, W., Lang, T., & Samuelsson, T. (2015). Evolution, diversification, and expression of KNOX proteins in plants. Frontiers in plant science, 6.
- Mukherjee, K., Brocchieri, L., & Bürglin, T. R. (2009). A comprehensive classification and evolutionary analysis of plant homeobox genes. Molecular biology and evolution, 26(12), 2775-2794.
- Bürglin, T. R., & Affolter, M. (2015). Homeodomain proteins: an update. Chromosoma, 1-25.
- Hofer, J., Gourlay, C., Michael, A., & Ellis, T. N. (2001). Expression of a class 1 knotted1-like homeobox gene is down-regulated in pea compound leaf primordia. Plant molecular biology, 45(4), 387-398.
- Imaichi, R., & Hiratsuka, R. (2007). Evolution of shoot apical meristem structures in vascular plants with respect to plasmodesmatal network. American journal of botany, 94(12), 1911-1921.
- Bharathan, G., Janssen, B. J., Kellogg, E. A., & Sinha, N. (1999). Phylogenetic relationships and evolution of the KNOTTED class of plant homeodomain proteins. Molecular Biology and Evolution, 16(4), 553-563.
- Bellaoui, M., Pidkowich, M. S., Samach, A., Kushalappa, K., Kohalmi, S. E., Modrusan, Z., ... & Haughn, G. W. (2001). The Arabidopsis BELL1 and KNOX TALE homeodomain proteins interact through a domain conserved between plants and animals. The Plant Cell, 13(11), 2455-2470
- Sinha, N. (1999). Leaf development in angiosperms. Annual review of plant biology, 50(1), 419-446.
- Hamant, O., & Pautot, V. (2010). Plant development: a TALE story. Comptes rendus biologies, 333(4), 371-381
- Janssen, B. J., Williams, A., Chen, J. J., Mathern, J., Hake, S., & Sinha, N. (1998). Isolation and characterization of two knotted-like homeobox genes from tomato. Plant molecular biology, 36(3), 417-425.
- Yu, H., Yang, S. H., & Goh, C. J. (2000). DOH1, a class 1 knox gene, is required for maintenance of the basic plant architecture and floral transition in orchid. The Plant Cell, 12(11), 2143-2159.