Cellules souches cancéreuses
Les cellules souches cancéreuses (CSCs) sont des cellules cancéreuses (présentes dans les tumeurs dites « solides » ou les cancers hématologiques) qui possèdent des caractéristiques associées aux cellules souches normales, notamment la capacité de donner naissance aux différentes populations de cellules présentes dans une tumeur particulière[1]. Les CSCs sont donc tumorigènes (formant des tumeurs), peut-être à la différence d'autres cellules cancéreuses non tumorigènes. Les CSCs peuvent générer des tumeurs grâce aux processus d'auto-renouvellement et de différenciation des cellules souches vers l’ensemble des composants cellulaires formant la tumeur. On suppose que ces cellules persistent dans les tumeurs comme une population distincte et provoquent des rechutes et des métastases en donnant naissance à de nouvelles tumeurs. Par conséquent, le développement de thérapies spécifiques ciblées sur les CSCs permet d'espérer une amélioration de la survie et de la qualité de vie de patients atteints de cancer, en particulier pour les patients atteints de maladies métastatiques.

Les traitements existants contre le cancer ont été principalement développés sur la base de modèles animaux, où les thérapies capables de favoriser la réduction des tumeurs ont été jugées efficaces. Cependant, les animaux ne reproduisent pas de manière fidèle la maladie humaine. En particulier, chez les souris, dont la durée de vie ne dépasse pas deux ans, la rechute des tumeurs est difficile à étudier.
L'efficacité des traitements anticancéreux est, dans les premiers stades des tests, correspond souvent à la fraction éliminée de cellules de la masse tumorale (fractional kill). Les CSCs ne représentant qu’une faible proportion de la masse tumorale, les traitements anticancéreux n’ont pas nécessairement d’effet spécifique sur elles. La théorie suggère que les chimiothérapies conventionnelles tuent les cellules différenciées ou en cours de différenciation, qui forment la masse de la tumeur mais qui ne génèrent pas de nouvelles cellules. Une population de CSCs, ayant donné naissance à la tumeur, pourrait rester intacte et provoquer une rechute.
Les CSCs ont été identifiées pour la première fois par John Dick dans la leucémie aiguë myéloïde à la fin des années 1990. Depuis le début des années 2000, elles font l'objet de recherches intensives en oncologie[2]. Le terme de CSC a été inventé par les biologistes Tannishtha Reya, Sean J. Morrison, Michael F. Clarke et Irving Weissman dans un article publié dans Nature en 2001[3].
Modèle de propagation tumorale
Dans différents sous-types de tumeurs, les cellules de la population tumorale présentent une hétérogénéité fonctionnelle et les tumeurs sont formées de cellules ayant des capacités de prolifération et de différenciation différentes[4]. Cette hétérogénéité fonctionnelle des cellules cancéreuses a conduit à la création de multiples modèles de propagation pour tenir compte de l'hétérogénéité et des différences de capacité de régénération des tumeurs : le modèle de cellule souche cancéreuse (CSC) et le modèle stochastique. Cependant, certaines études maintiennent que cette différence est artificielle, puisque les deux processus agissent de manière complémentaire en ce qui concerne les populations tumorales réelles[1].
Modèle cellule souche cancéreuse
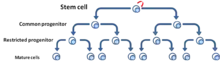
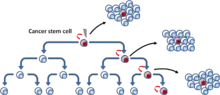
Le modèle de CSC, également connu sous le nom de modèle hiérarchique, propose que les tumeurs soient organisées de manière hiérarchique (les CSCs se trouvant au sommet[5] (Fig. 3)). Dans la population des tumeurs, il existe des CSCs qui sont des cellules tumorigènes et qui sont biologiquement distinctes des autres sous-populations[6]. Elles présentent deux caractéristiques déterminantes : leur capacité à s'auto-renouveler indéfiniment et leur capacité à se différencier en une descendance non tumorigène mais qui contribue quand même à la croissance de la tumeur. Ce modèle suggère que seules certaines sous-populations de CSCs ont la capacité d’induire la progression tumorale, ce qui signifie qu'il existe des caractéristiques spécifiques (intrinsèques) qui peuvent être identifiées et ensuite ciblées pour détruire une tumeur à long terme sans qu'il soit nécessaire de combattre la tumeur dans son ensemble[7].
Modèle stochastique
Selon le "modèle stochastique" (ou "modèle d'évolution clonale"), chaque cellule cancéreuse d'une tumeur pourrait acquérir la capacité de s'auto-renouveler et de se différencier en fonction des lignées nombreuses et hétérogènes de cellules cancéreuses qui compromettent une tumeur[8]. Ainsi, pour qu'une cellule devienne cancéreuse, elle doit subir un nombre important d'altérations de sa séquence d'ADN. Ce modèle cellulaire suggère que ces mutations pourraient se produire dans n'importe quelle cellule du corps et provoquer un cancer. Essentiellement, cette théorie propose que toutes les cellules possèdent la capacité d'être tumorigènes, rendant toutes les cellules tumorales équipotentes avec la capacité de s'auto-renouveler ou de se différencier, conduisant à l'hétérogénéité de la tumeur tandis que d'autres peuvent se différencier en cellules non-CSCs[6] - [9]. Le potentiel de la cellule peut être influencé par des facteurs génétiques ou épigénétiques (mutations), entraînant une diversité phénotypique des cellules tumorigènes et non-tumorigènes qui composent la tumeur.
Ces mutations pourraient s'accumuler progressivement et améliorer la résistance et l'aptitude des cellules qui leur permettent de concurrencer d'autres cellules tumorales, mieux connu sous le nom de modèle d'évolution somatique[6]. Le modèle d'évolution clonale, qui se produit à la fois dans le modèle CSC et le modèle stochastique, postule que les cellules tumorales mutantes, ayant un avantage de croissance, surpassent les autres. Les cellules de la population dominante ont un potentiel similaire pour initier la croissance d'une tumeur[10]. (Fig. 4).
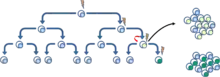
[11] Ces deux modèles ne sont pas mutuellement exclusifs, car les CSCs elles-mêmes subissent une évolution clonale. Ainsi, d’autres CSCs peuvent émerger si une mutation leur confère des propriétés plus agressives[12] (Fig. 5).
Relation modèle CSC et modèle stochastique
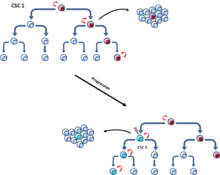
Une étude réalisée en 2014 soutient que le fossé entre ces deux modèles controversés peut être comblé en fournissant une explication alternative sur l'hétérogénéité des tumeurs. Les chercheurs ont démontré un modèle qui inclut à la fois des aspects du modèle stochastique et du modèle CSC[9]. Ils ont examiné la plasticité des cellules souches cancéreuses, dans laquelle les cellules souches cancéreuses peuvent faire la transition entre les cellules souches non cancéreuses (non-CSCs) et les CSCs in situ, ce qui soutient un modèle plus stochastique[9] - [13]. Toutefois, l'existence de populations biologiquement distinctes de non-CSC et de CSC soutient un modèle plus CSC, ce qui laisse entendre que les deux modèles peuvent jouer un rôle vital dans l'hétérogénéité des tumeurs[9].
Modèle immunologique CSC
Ce modèle suggère que les propriétés immunologiques peuvent être importantes pour comprendre la tumorigenèse et l'hétérogénéité. Ainsi, les CSCs peuvent être très rares dans certaines tumeurs[14], mais certains chercheurs ont découvert qu'une grande proportion de cellules tumorales peuvent initier des tumeurs si elles sont transplantées dans des souris immunodéprimées[15]. Ces données remettent en question la pertinence d’une population rare de CSCs. Cependant, les cellules souches[16] et les CSCs[17] possèdent des propriétés immunologiques uniques qui les rendent plus résistantes à l'immunosurveillance. Alors, seules les CSCs peuvent être capables d'ensemencer des tumeurs chez des patients faisant l'objet d'une immunosurveillance fonctionnelle, et le privilège immunitaire peut être un critère clé pour identifier les CSCs[18]. En outre, le modèle suggère que les CSCs peuvent initialement dépendre de niches de cellules souches, et les CSCs peuvent y fonctionner comme un réservoir dans lequel des mutations peuvent s'accumuler pendant des décennies sans être limitées par le système immunitaire. Des tumeurs peuvent spontanément se développer si: A) les CSCs perdent leur dépendance vis-à-vis des niches de CS (tumeurs moins différenciées), B) leur progéniture de cellules tumorales normales hautement prolifératives, mais initialement immunogènes, développe des moyens d'échapper à l'immunosurveillance ou, C) le système immunitaire peut perdre sa capacité de suppression des tumeurs, par exemple en raison du vieillissement[18].
Débat
L'existence des CSCs fait l'objet de débats, car de nombreuses études n'ont trouvé aucune cellule présentant leurs caractéristiques spécifiques[14]. Les cellules cancéreuses doivent être capables de proliférer et de se renouveler en permanence afin de conserver les nombreuses mutations nécessaires à la cancérogenèse et soutenir la croissance d'une tumeur. En effet, les cellules différenciées (limitées par la limite de Hayflick[19]) ne peuvent pas se diviser indéfiniment. Sur le plan thérapeutique, si la plupart des cellules tumorales sont dotées de propriétés de cellules souches, cibler directement la taille de la tumeur est une stratégie valable. Si les CSCs représentent une petite minorité, les cibler peut être plus efficace. Un autre débat porte sur l'origine des CSCs - qu'il s'agisse de la dérégulation des cellules souches normales ou d'une population plus spécialisée qui a acquis la capacité de se renouveler elle-même (ce qui est lié à la question de la plasticité des cellules souches). La découverte d’une plasticité phénotypique des cellules cancéreuses induite par la thérapie, altérant leurs transcriptomes et leur conférant des propriétés de type cellule souche pour échapper à la destruction, vient troubler ce débat[20].
Mise en évidence
Les premières preuves concluantes concernant l’existence des CSC sont apparues en 1997. Bonnet et Dick ont isolé une sous-population de cellules leucémiques qui exprimaient le marqueur de surface CD34, mais pas CD38[21]. Les auteurs ont établi que la sous-population CD34+/CD38− est capable d'initier des tumeurs chez les souris NOD/SCID qui étaient histologiquement similaires au donneur. La première preuve de l'existence de cellule souche cancéreuse dans une tumeur solide a suivi en 2002 avec la découverte d'un clonogène, une cellule sphérique isolée et caractérisée à partir de gliomes de cerveau humain adulte. Les tumeurs gliales corticales humaines contiennent des cellules souches neurales exprimant des marqueurs astrogliaux et neuronaux in vitro[22]. Il a été démontré que les cellules souches cancéreuses isolées de gliomes humains adultes induisent des tumeurs qui ressemblent à la tumeur mère lorsqu'elles sont greffées sur des modèles intracrâniens de souris nude[23].
Dans les expériences de recherche sur le cancer, des cellules tumorales sont parfois injectées à un animal de laboratoire pour établir une tumeur. La progression de la maladie est alors suivie dans le temps et l'efficacité des nouveaux médicaments peut être testée. La formation d'une tumeur nécessite l'introduction de milliers ou de dizaines de milliers de cellules. Classiquement, cela s'expliquait par une mauvaise méthodologie (c'est-à-dire que les cellules tumorales perdent leur viabilité lors du transfert) ou par l'importance critique du micro-environnement, l'environnement biochimique particulier des cellules injectées. Les partisans du paradigme des CSC affirment que seule une petite fraction des cellules injectées, les CSC, a le potentiel de générer une tumeur. Dans la leucémie aiguë myéloïde humaine, la fréquence de ces cellules est inférieure à 1 sur 10 000[21].
D'autres preuves proviennent de l'histologie. De nombreuses tumeurs sont hétérogènes et contiennent de multiples types de cellules natives de l'organe hôte. L'hétérogénéité de la tumeur est généralement conservée par les métastases de la tumeur. Cela suggère que la cellule qui les a produites avait la capacité de générer de multiples types de cellules, une caractéristique classique des cellules souches[21].
L'existence des cellules souches de la leucémie a incité la recherche sur d'autres cancers. Des CSC ont été identifiées dans plusieurs tumeurs solides, notamment:
- Cerveau[24]
- Sein[25]
- Côlon[26]
- Ovaire[27] - [28]
- Pancréas[29]
- Prostate[30] - [31]
- Mélanome[32] - [33] - [34] - [35]
- Myélome Multiple[36] - [37]
- Cancer de la peau non mélanome[38] - [39]
Modèles mécanique et mathématique
Après avoir émis des hypothèses sur les mécanismes conduisant au cancer, il est possible de développer des modèles mathématiques prédictifs[40] en se basant sur la méthode du compartiment cellulaire. Par exemple, les croissances de cellules anormales peuvent être désignées par des probabilités de mutations spécifiques. Un tel modèle prédit qu’une succession d’altérations génétiques des cellules matures augmente la formation d'une progéniture anormale et le risque de cancer[41]. L'efficacité clinique de tels modèles[42] reste encore à établir.
Origine

L'origine des CSCs est un domaine de recherche actif. La réponse peut dépendre du type de tumeur et du phénotype. Jusqu'à présent, l'hypothèse selon laquelle les tumeurs proviennent d'une seule "cellule d'origine" n'a pas été démontrée à l'aide du modèle des cellules souches cancéreuses. Cela s'explique par le fait que les cellules souches cancéreuses ne sont pas présentes dans les tumeurs en phase terminale.
Les hypothèses sur l’origine des CSCs comprennent des mutants dans des cellules souches progénitrices ou en développement, des mutants dans des cellules souches adultes ou des cellules progénitrices adultes et des mutants dans des cellules différenciées qui acquièrent des attributs de type souches. Ces théories se concentrent souvent sur la "cellule d'origine" d'une tumeur.
Mutation des cellules souches
L'hypothèse de la "mutation des populations de niche de cellules souches au cours du développement" affirme que ces populations de cellules souches en développement subissent une mutation puis se multiplient de sorte que la mutation est partagée par de nombreux descendants. Ces cellules filles sont beaucoup plus proches de devenir des tumeurs et leur nombre augmente la probabilité d'une mutation cancéreuse[43].
Cellules souches adultes
Une autre théorie associe les cellules souches adultes (CSA) à la formation de tumeurs. Cette dernière est le plus souvent associée à des tissus présentant un taux élevé de renouvellement cellulaire (comme la peau ou l'intestin). Dans ces tissus, les CSA représentent des cibles idéales en raison de leurs fréquentes divisions cellulaires (par rapport à la plupart des CSA) et de leur longue durée de vie. Cette combinaison crée un ensemble de circonstances favorables à l'accumulation de mutations : l'accumulation de mutations est le principal facteur qui entraîne l'apparition du cancer. Les preuves montrent que cette association représente un phénomène réel, bien que des cancers spécifiques aient été liés à une cause précise[44] - [45].
Dé-différenciation
La dé-différenciation des cellules mutées peut créer des caractéristiques semblables à celles des cellules souches, ce qui suggère que n'importe quelle cellule peut devenir une cellule souche cancéreuse. En d'autres termes, une cellule entièrement différenciée subit des mutations ou des signaux extracellulaires qui la ramènent à un état de cellule souche. Ce concept a été démontré tout récemment dans des modèles de cancer de la prostate. Le traitement par privation androgénique semble altérer transitoirement le transcriptome des cellules en cellule souche de type crête neurale, avec les propriétés invasives et multipotentes de cette classe de cellules souches[20].
Hiérarchie
Le concept de hiérarchie des tumeurs affirme qu'une tumeur est une population hétérogène de cellules mutantes, qui partagent toutes certaines mutations, mais varient en fonction d'un phénotype spécifique. Une tumeur héberge plusieurs types de cellules souches, des cellules présentant un avantage sélectif pour l'environnement spécifique et d'autres cellules moins prospères. Ces lignées secondaires peuvent être plus avantagées dans d'autres environnements, ce qui permet à la tumeur de s'adapter, y compris à une intervention thérapeutique. Si ce concept est correct, il a une incidence sur les régimes de traitement spécifiques des cellules souches cancéreuses[46]. Une telle hiérarchie compliquerait les tentatives visant à en déterminer l'origine.
Identification
Les CSCs, qui sont maintenant signalées dans la plupart des tumeurs humaines, sont couramment identifiées et enrichies à l'aide de stratégies d'identification des cellules souches normales qui sont similaires d'une étude à l'autre[47]. Ces procédures comprennent le tri des cellules activées par fluorescence (FACS), avec des anticorps dirigés contre les marqueurs de surface cellulaire et des approches fonctionnelles comprenant un test permettant d’isoler une population cellulaire appelée "side population » ou un test Aldefluor[48]. Les populations enrichies en CSCs sont ensuite implantées, à différentes doses, chez des souris immunodéficientes pour évaluer leur capacité de développement tumoral. Ce test in vivo est appelé test de dilution limite. Les sous-ensembles de cellules tumorales qui peuvent initier le développement de la tumeur à un faible nombre de cellules sont ensuite testés pour leur capacité d'auto-renouvellement dans des études tumorales en série[49].
Les CSCs peuvent également être identifiées par une coloration Hoechst. Cette technique est basée sur l’efflux du colorant via des transporteurs membranaires de multiples drogues (MDR) et de la famille ABC (ATP binding cassettes)[48].
Une autre approche consiste à effectuer des tests de sphères. De nombreuses cellules souches normales, comme les cellules hématopoïétiques ou les cellules souches de tissus, dans des conditions de culture spéciales, forment des sphères tridimensionnelles qui peuvent se différencier. Comme les cellules souches normales, les CSCs isolées de tumeurs du cerveau ou de la prostate ont également la capacité de former des sphères indépendantes d'ancrage[50].
Hétérogénéité (marqueurs)
Des CSCs ont été identifiées dans diverses tumeurs solides. Généralement, des marqueurs spécifiques aux cellules souches normales sont utilisés pour isoler les CSCs des tumeurs solides et hématologiques. Les marqueurs les plus fréquemment utilisés pour l'isolement des CSCs sont les suivants : CD133 (également appelé PROM1), CD44, ALDH1A1, CD34, CD24 et EpCAM (molécule d'adhésion des cellules épithéliales, également appelée antigène spécifique de l'épithélium, ESA)[51].
CD133 (prominine 1) est une glycoprotéine à cinq domaines transmembranaires exprimée par les cellules souches et progénitrices CD34+, dans les précurseurs endothéliaux et les cellules souches neurales fœtales. Elle a été détectée grâce à son épitope glycosylé appelé AC133.
EpCAM (molécule d'adhésion des cellules épithéliales , ESA, TROP1) est une molécule d'adhésion cellulaire hémophile indépendante de Ca2+, exprimée sur la surface basolatérale de la plupart des cellules épithéliales.
CD90 (THY1) est une glycosylphosphatidylinositol glycoprotéine ancrée dans la membrane plasmique et impliquée dans la transduction du signal. Elle peut également jouer un rôle de médiateur dans l'adhésion entre les thymocytes et le stroma thymique.
CD44 (PGP1) est une molécule d'adhésion qui joue un rôle pléiotropique dans la signalisation, la migration et le guidage des cellules. Elle a de multiples isoformes, dont CD44H, qui présente une grande affinité pour l'hyaluronate, et CD44V, qui a des propriétés métastatiques.
CD24 (HSA) est une molécule d'adhésion glycosylée fixée au glycosylphosphatidylinositol , qui joue un rôle de co-stimulation dans les cellules B et T.
CD200 (OX-2) est une glycoprotéine membranaire de type 1, qui délivre un signal inhibiteur aux cellules immunitaires, dont les cellules T, les cellules tueuses naturelles (NK, natural killer) et les macrophages.
Les ALDH sont une famille d'enzymes omniprésentes de la famille des aldéhydes déshydrogénases, qui catalysent l'oxydation des aldéhydes aromatiques en acides carboxyliques. Par exemple, elles jouent un rôle dans la conversion du rétinol en acide rétinoïque, qui est essentiel à la survie[52] - [53].
La première tumeur maligne solide à partir de laquelle les CSCs ont été isolées et identifiées était le cancer du sein. Les CSCs issues de cancer du sein sont les plus étudiées. Les CSCs du sein ont été enrichies en sous-populations CD44+CD24-/low[25], SP[54] et ALDH+[55] - [56]. Les CSCs du sein présentent donc des phénotypes divers. L'expression des marqueurs des CSCs dans les cellules de cancer du sein est apparemment hétérogène et les populations de CSCs du sein varient selon les tumeurs[57]. Les populations de cellules CD44+CD24− et CD44+CD24+ sont toutes deux des cellules initiatrices de tumeurs ; cependant, les CSCs sont plus fortement enrichies en utilisant le profil de marqueur CD44+CD49fhiCD133/2hi[58].
Des CSCs ont été identifiées dans de nombreuses tumeurs cérébrales. Les cellules tumorales de type souches ont été identifiées à l'aide de marqueurs de surface cellulaire, notamment CD133[59], SSEA-1 (stage-specific embryonic antigen-1)[60], EGFR[61] et CD44[62]. Dans les tumeurs cérébrales, l'utilisation de CD133 pour l'identification de cellules souches peut être problématique. D’une part, les cellules tumorigènes se trouvent à la fois dans les populations de cellules CD133+ et CD133− dans certains gliomes. D’autre part, certaines cellules CD133+ peuvent ne pas posséder de capacité d'initiation tumorale[61].
Des CSCs ont été décrites dans le cancer du côlon[26]. Pour les identifier, des marqueurs de surface cellulaire tels que CD133[26], CD44[63] et ABCB5[64], l'analyse fonctionnelle, ainsi que l'analyse clonale[65] et le test Aldefluor ont été utilisés[66]. L'utilisation de CD133 comme marqueur positif pour les CSCs du côlon a donné des résultats contradictoires. L'épitope AC133, mais pas la protéine CD133, est spécifiquement exprimé dans les CSCs du côlon et son expression est perdue lors de la différenciation[67]. En outre, les cellules CD44+ du cancer du côlon et une sous-fraction supplémentaire de la population de cellules CD44+EpCAM+ avec CD166 améliorent le succès des greffes de tumeurs[63].
De multiples CSCs ont été signalées dans la prostate[68], les poumons et de nombreux autres organes, notamment le foie, le pancréas, les reins ou les ovaires[52] - [69]. Dans le cancer de la prostate, les cellules initiatrices de tumeur ont été identifiées dans le sous-ensemble de cellules CD44+[70] comme étant des cellules CD44+α2β1+[71], TRA-1-60+CD151+CD166+ [72] ou ALDH+[73]. Des marqueurs putatifs pour les CSCs pulmonaires ont été décrits, notamment CD133+[74], ALDH+[75], CD44+ [76] et la protéine onco-fœtale 5T4+[77].
Métastases
Les métastases sont la cause principale de la létalité des tumeurs. Cependant, toutes les cellules tumorales ne peuvent pas métastaser. Ce potentiel dépend de facteurs qui déterminent la croissance, l'angiogenèse, l'invasion et d'autres processus fondamentaux.
Transition épithélio-mésenchymateuse
Dans les tumeurs épithéliales, la transition épithélio-mésenchymateuse (TEM) est considérée comme un événement crucial[78]. La TEM et la transition inverse, à savoir la transition mésenchymateuse-épithéliale (TME), sont impliquées dans le développement embryonnaire, qui implique la perturbation de l'homéostasie des cellules épithéliales et l'acquisition d'un phénotype mésenchymateux migrateur[79]. La TEM semble être contrôlée par des voies canoniques telles que la voie WNT et la voie du TGFb (le facteur de croissance transformant β)[80].
Une caractéristique importante de la TEM est la perte de l’expression membranaire de la E-cadhérine au niveau des jonctions adhérentes, où la β-caténine pourrait jouer un rôle important. La translocation de la ß-caténine des jonctions adhérentes vers le noyau peut entraîner une perte de la E-cadhérine et, par conséquent, une TEM. La β-caténine nucléaire peut apparemment activer directement, par transcription, les gènes cibles associés à la TEM, tels que SLUG (également connu sous le nom de SNAI2), répresseur transcriptionnel du gène codant la E-cadhérine[81]. Les propriétés mécaniques du microenvironnement de la tumeur, comme l'hypoxie, peuvent contribuer à la survie et au potentiel métastatique de la CSC par la stabilisation des facteurs induits par l'hypoxie grâce aux interactions avec les dérivés réactifs de l’oxygène (ROS, reactive oxygen species)[82] - [83].
Les cellules tumorales soumises à une TEM peuvent être des précurseurs de cellules cancéreuses métastatiques, voire de CSCs métastatiques[84]. Dans la bordure invasive du carcinome pancréatique, un sous-ensemble de cellules CD133+CXCR4+ (récepteur de la chimiokine CXCL12 également connu sous le nom de ligand SDF1) a été défini. Ces cellules présentaient une activité migratoire nettement plus forte que leurs homologues CD133+CXCR4−, mais toutes deux avaient une capacité de développement tumoral similaire[85]. De plus, l'inhibition du récepteur CXCR4 a réduit le potentiel métastatique sans altérer la capacité tumorigène[86].
Motif d’expression en deux phases
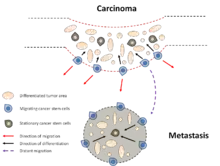
Dans le cancer du sein, les cellules CD44+CD24−/low sont détectables dans les épanchements pleuraux métastatiques[25]. En revanche, un nombre accru de cellules CD24+ a été identifié dans les métastases distantes chez les patientes atteintes d'un cancer du sein[87]. Il est possible que les cellules CD44+CD24−/low se détachent de la tumeur initiale pour former des métastases et une fois dans leur nouveau site, elles changent de phénotype et subissent une différenciation limitée[88]. L'hypothèse du schéma d'expression en deux phases propose deux formes de cellules souches cancéreuses : stationnaires (CSS) et mobiles (CSM). Les CSS sont ancrées dans les tissus et persistent dans les zones différenciées tout au long de la progression de la tumeur. Les CSM sont situées à l'interface tumeur-hôte. Ces cellules sont apparemment dérivées des CSS à travers l'acquisition d'une TEM transitoire (Fig. 7)[89].
Applications
Le ciblage des CSCs jouent un rôle dans le traitement contre le cancer, notamment à travers l'identification de la maladie, la sélection de médicaments cibles, la prévention des métastases et les stratégies d'intervention.
Traitement
Les CSCs sont intrinsèquement plus résistantes aux agents chimiothérapeutiques. Cinq facteurs principaux y contribuent[90] :
- 1. Leur niche les protège contre le contact avec de grandes concentrations de médicaments anticancéreux;
- 2. Elles expriment diverses protéines transmembranaires, telles que MDR1 et BCRP, qui pompent les médicaments hors du cytoplasme;
- 3. Elles se divisent lentement, comme les cellules souches adultes ont tendance à le faire, et ne sont donc pas tuées par les agents chimiothérapeutiques qui ciblent les cellules qui se répliquent rapidement en endommageant l'ADN ou en inhibant la mitose;
- 4. Elles régulent les protéines de réparation des dommages à l'ADN;
- 5. Elles sont caractérisées par une suractivation des voies de signalisation anti-apoptotiques.
Après une chimiothérapie, les CSCs ayant résisté au traitement sont capables de repeupler la tumeur et de provoquer une rechute. Un traitement supplémentaire visant à éliminer les CSCs en plus des cellules somatiques cancéreuses doit être utilisé pour empêcher cela.
Ciblage (Thérapie ciblée)
Le ciblage sélectif des CSCs peut permettre de traiter des tumeurs agressives et non résécables, ainsi que de prévenir les métastases et les rechutes[91]. L'hypothèse suggère que lors de l'élimination des CSCs, le cancer pourrait régresser en raison de la différenciation et/ou de la mort cellulaire. La fraction des cellules tumorales qui sont des CSCs et qui doivent donc être éliminées reste floue[92].
Des études ont cherché à identifier des marqueurs spécifiques[25] et des signatures tumorales protéomiques et génomiques qui distinguent les CSCs des autres cellules[93]. En 2009, une équipe de scientifiques a montré que la salinomycine pouvait réduire de façon sélective et par plus de 100 fois la proportion de CSCs du sein chez des souris par rapport au Paclitaxel, un agent chimiothérapeutique couramment utilisé[94]. Certains types de cellules cancéreuses peuvent survivre au traitement à la salinomycine par autophagie[95]. C'est-à-dire que les cellules utilisent des organites acides tels que les lysosomes pour dégrader et recycler certains types de protéines. L'utilisation d'inhibiteurs de l’autophagie peut tuer les cellules souches cancéreuses qui survivent grâce à ce processus[96].
Le récepteur de surface cellulaire interleukine-3-alpha (CD123) est surexprimé sur les cellules souches leucémiques (CSL) CD34+CD38− dans la leucémie aiguë myéloblastique (LAM) mais pas sur les cellules normales CD34+CD38− de la moelle osseuse[97]. Le traitement de souris NOD/SCID ayant subi une greffe de LAM, par un anticorps monoclonal anti-CD123 altère le foyer de CSLs au niveau de la moelle osseuse et réduit la prolifération globale des cellules de LAM ainsi que la proportion de CSLs chez les souris receveuses[98].
Une étude réalisée en 2015 a permis de conditionner des nanoparticules avec du miR-34a et du bicarbonate d'ammonium et de les administrer à des CSCs de prostate dans un modèle de souris. Ensuite, les chercheurs ont irradié la zone avec une lumière laser proche de l’infrarouge. Ceci conduit à un gonflement d’au moins trois fois plus la taille des nanoparticules, faisant éclater les endosomes et dispersant l'ARN dans la cellule. Le miR-34a peut faire baisser les niveaux de CD44[99] - [100].
Une étude de 2018 a identifié des inhibiteurs de la famille des enzymes ALDH1A et a montré qu'ils pouvaient dépléter sélectivement les cellules souches cancéreuses putatives dans plusieurs lignées cellulaires de cancer de l'ovaire[101].
Voies de signalisation
Le développement de nouveaux médicaments ciblant les CSCs nécessite une meilleure compréhension des mécanismes cellulaires qui régulent la prolifération cellulaire. Les premiers progrès dans ce domaine ont été réalisés avec les cellules souches hématopoïétiques (CSH) et leurs homologues transformés dans la leucémie, la maladie pour laquelle l'origine des CSCs est la mieux comprise. Les cellules souches de nombreux organes partagent les mêmes voies cellulaires que les CSHs issues de leucémies.
Une cellule souche normale peut être transformée en CSC en dérégulant les voies de prolifération et de différenciation qui la contrôlent ou en induisant une activité oncoprotéique.
BMI-1
Le répresseur transcriptionnel du groupe Polycomb, Bmi-1, a été découvert comme un oncogène commun activé dans les lymphomes[102] et a ensuite été montré comme régulant les CSHs[103]. Le rôle de Bmi-1 a été illustré dans les cellules souches neurales[104]. La voie semble être active dans les CSCs des tumeurs cérébrales pédiatriques[105].
Notch
La voie Notch joue un rôle dans le contrôle de la prolifération des cellules souches pour plusieurs types de cellules, notamment les CS hématopoïétiques, neurales et mammaires[106]. Des composants de cette voie ont été proposés pour agir comme oncogènes dans les tumeurs mammaires[107] et autres.
Une branche de la voie de signalisation Notch qui implique le facteur de transcription Hes3 régule en culture le nombre de cellules de glioblastome qui présentent les caractéristiques de CSCs[108].
Sonic hedgehog et Wnt
Ces voies de développement sont des régulateurs des cellules souches (CS)[109]. Les voies Sonic hedgehog (SHH) et Wnt sont toutes deux couramment hyperactivées dans les tumeurs et sont nécessaires pour soutenir la croissance tumorale. Toutefois, les facteurs de transcription Gli qui sont régulés par SHH tirent leur nom des gliomes, où ils sont fortement exprimés. Il existe un certain degré d’interférence entre les deux voies et elles sont généralement activées ensemble[110]. En revanche, dans le cancer du côlon, la voie de signalisation Hedgehog semble antagoniser la voie Wnt[111].
Il existe des inhibiteurs de la voie Sonic hedgehog, comme la cyclopamine. La cyclopamine sous sa forme soluble peut être plus efficace dans le traitement du cancer. Le diméthylaminoparthénolide (DMAPT), un dérivé hydrosoluble du parthénolide, induit un stress oxydatif et inhibe la signalisation NF-κB[112] dans la LAM (leucémie) et éventuellement dans le myélome et le cancer de la prostate. La télomérase est un sujet d'étude dans la physiologie des CSCs[113]. Récemment, un essai clinique évaluant l’effet du GRN163L (Imetelstat) sur le ciblage des cellules souches du myélome a débuté.
La signalisation Wnt peut devenir indépendante de stimuli réguliers, à travers des mutations dans les oncogènes en aval et les gènes suppresseurs de tumeurs qui sont activés de façon permanente même si le récepteur normal n'a pas reçu de signal. La β-caténine se lie à des facteurs de transcription tels que la protéine TCF4 et, en combinaison, les molécules activent les gènes nécessaires. Le LF3 inhibe fortement la liaison de la ß-caténine in vitro, dans les lignées cellulaires et réduit la croissance des tumeurs dans des modèles de souris. Il empêche la réplication et réduit leur capacité à migrer, le tout sans affecter les cellules saines. Après le traitement par le LF3, il ne restait plus de cellules souches cancéreuses. Cette découverte est le fruit d'une "conception rationnelle des médicaments", faisant appel aux technologies AlphaScreens et ELISA[114].
La voie Wnt/ß-caténine est régulièrement activée dans les cellules cancéreuses conduisant à l’expression de gènes cible de façon constitutive[115]. L’un des gènes activés, GAST, code hPG80 ou progastrine circulante dans le sang. Initialement considérée comme biologiquement inactive, la hPG80 s'est avérée participer à certaines caractéristiques d'une tumeur, telles que la perturbation des jonctions cellule-cellule[116], la prolifération cellulaire[117], l'inhibition de l'apoptose[118], la régulation des cellules souches cancéreuses[119] - [120] et l'angiogenèse[121]. En outre, il a été démontré que le ciblage de la hPG80 par un anticorps spécifique favorise l'apoptose, diminue la prolifération et la migration des cellules cancéreuses colorectales humaines, et inhibe l'auto-renouvellement des CSCs de différentes origines, ainsi que la tumorigenèse Wnt chez la souris[119]. Plus récemment il a été démontré que la hPG80 était exprimée par différents types de cancers même à des stades précoces[122].
Notes et références
Notes
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Cancer Stem Cell » (voir la liste des auteurs).
Références
- Sreepadmanabh M, « Investigations into the cancer stem cell niche using in-vitro 3-D tumor models and microfluidics », Biotechnology Advances, vol. 36, no 4, , p. 1094–1110 (PMID 29559382, DOI 10.1016/j.biotechadv.2018.03.009)
- Siddhartha Mukherjee, « The Cancer Sleeper Cell », New York Times, (lire en ligne, consulté le )
- (en) Tannishtha Reya, Sean J. Morrison, Michael F. Clarke et Irving L. Weissman, « Stem cells, cancer, and cancer stem cells », Nature, vol. 414, no 6859, , p. 105–111 (ISSN 0028-0836, PMID 11689955, DOI 10.1038/35102167, hdl 2027.42/62862)
- Heppner GH, Miller BE, « Tumor heterogeneity: biological implications and therapeutic consequences », Cancer Metastasis Reviews, vol. 2, no 1, , p. 5–23 (PMID 6616442, DOI 10.1007/BF00046903)
- Bonnet D, Dick JE, « Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell », Nature Medicine, vol. 3, no 7, , p. 730–7 (PMID 9212098, DOI 10.1038/nm0797-730)
- (en) Benjamin Beck et Cédric Blanpain, « Unravelling cancer stem cell potential », Nature Reviews Cancer, vol. 13, no 10, , p. 727–738 (ISSN 1474-1768, DOI 10.1038/nrc3597, lire en ligne, consulté le )
- Shackleton M, Quintana E, Fearon ER, Morrison SJ, « Heterogeneity in cancer: cancer stem cells versus clonal evolution », Cell, vol. 138, no 5, , p. 822–9 (PMID 19737509, DOI 10.1016/j.cell.2009.08.017)
- (en) Antonija Kreso et John E. Dick, « Evolution of the Cancer Stem Cell Model », Cell Stem Cell, vol. 14, no 3, , p. 275–291 (ISSN 1934-5909 et 1875-9777, PMID 24607403, DOI 10.1016/j.stem.2014.02.006, lire en ligne, consulté le )
- (en) Weikang Wang, Yi Quan, Qibin Fu et Yu Liu, « Dynamics between Cancer Cell Subpopulations Reveals a Model Coordinating with Both Hierarchical and Stochastic Concepts », PLOS ONE, vol. 9, no 1, , e84654 (ISSN 1932-6203, PMID 24416258, PMCID PMC3886990, DOI 10.1371/journal.pone.0084654, lire en ligne, consulté le )
- Barabé F, Kennedy JA, Hope KJ, Dick JE, « Modeling the initiation and progression of human acute leukemia in mice », Science, vol. 316, no 5824, , p. 600–4 (PMID 17463288, DOI 10.1126/science.1139851, Bibcode 2007Sci...316..600B)
- Nowell PC, « The clonal evolution of tumor cell populations. », Science, vol. 194, no 4260, , p. 23–8 (PMID 959840, DOI 10.1126/science.959840, Bibcode 1976Sci...194...23N)
- Clark EA, Golub TR, Lander ES, Hynes RO, « Genomic analysis of metastasis reveals an essential role for RhoC », Nature, vol. 406, no 6795, , p. 532–5 (PMID 10952316, DOI 10.1038/35020106)
- (en) Marina Carla Cabrera, Robert E. Hollingsworth et Elaine M. Hurt, « Cancer stem cell plasticity and tumor hierarchy », World Journal of Stem Cells, vol. 7, no 1, , p. 27–36 (PMID 25621103, PMCID PMC4300934, DOI 10.4252/wjsc.v7.i1.27, lire en ligne, consulté le )
- (en) Piyush B. Gupta, Christine L. Chaffer et Robert A. Weinberg, « Cancer stem cells: mirage or reality? », Nature Medicine, vol. 15, no 9, , p. 1010–1012 (ISSN 1546-170X, DOI 10.1038/nm0909-1010, lire en ligne, consulté le )
- E Quintana, M Shackleton, M. S Sabel, D. R Fullen, T. M Johnson et S. J Morrison, « Efficient tumour formation by single human melanoma cells », Nature, vol. 456, no 7222, , p. 593–8 (PMID 19052619, PMCID 2597380, DOI 10.1038/nature07567, Bibcode 2008Natur.456..593Q)
- Ichiryu N, Fairchild PJ, Embryonic Stem Cell Immunobiology, vol. 1029, coll. « Methods in Molecular Biology », , 1–16 p. (ISBN 978-1-62703-477-7, PMID 23756938, DOI 10.1007/978-1-62703-478-4_1), « Immune Privilege of Stem Cells »
- C MacCalli, A Volontè, C Cimminiello et G Parmiani, « Immunology of cancer stem cells in solid tumours. A review », European Journal of Cancer, vol. 50, no 3, , p. 649–55 (PMID 24333096, DOI 10.1016/j.ejca.2013.11.014)
- V. S Bruttel et J Wischhusen, « Cancer stem cell immunology: Key to understanding tumorigenesis and tumor immune escape? », Frontiers in Immunology, vol. 5, , p. 360 (PMID 25120546, PMCID 4114188, DOI 10.3389/fimmu.2014.00360)
- Hayflick L, « The limited in vitro lifetime of human diploid cell strains », Experimental Cell Research, vol. 37, no 3, , p. 614–36 (PMID 14315085, DOI 10.1016/0014-4827(65)90211-9)
- Nouri M, Caradec J, Lubik AA, Li N, Hollier BG, Takhar M, Altimirano-Dimas M, Chen M, Roshan-Moniri M, Butler M, Lehman M, Bishop J, Truong S, Huang SC, Cochrane D, Cox M, Collins C, Gleave M, Erho N, Alshalafa M, Davicioni E, Nelson C, Gregory-Evans S, Karnes RJ, Jenkins RB, Klein EA, Buttyan R, « Therapy-induced developmental reprogramming of prostate cancer cells and acquired therapy resistance », Oncotarget, vol. 8, no 12, , p. 18949–18967 (PMID 28145883, PMCID 5386661, DOI 10.18632/oncotarget.14850)
- Bonnet D, Dick JE, « Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell », Nature Medicine, vol. 3, no 7, , p. 730–7 (PMID 9212098, DOI 10.1038/nm0797-730)
- Ignatova TN, Kukekov VG, Laywell ED, Suslov ON, Vrionis FD, Steindler DA, « Human cortical glial tumors contain neural stem-like cells expressing astroglial and neuronal markers in vitro », Glia, vol. 39, no 3, , p. 193–206 (PMID 12203386, DOI 10.1002/glia.10094)
- Sundar SJ, Hsieh JK, Manjila S, Lathia JD, Sloan A, « The role of cancer stem cells in glioblastoma », Neurosurgical Focus, vol. 37, no 6, , E6 (PMID 25434391, DOI 10.3171/2014.9.FOCUS14494).
- Singh SK, Clarke ID, Terasaki M, Bonn VE, Hawkins C, Squire J, Dirks PB, « Identification of a cancer stem cell in human brain tumors », Cancer Research, vol. 63, no 18, , p. 5821–8 (PMID 14522905, lire en ligne).
- Al-Hajj M, Wicha MS, Benito-Hernandez A, Morrison SJ, Clarke MF, « Prospective identification of tumorigenic breast cancer cells », Proceedings of the National Academy of Sciences of the United States of America, vol. 100, no 7, , p. 3983–8 (PMID 12629218, PMCID 153034, DOI 10.1073/pnas.0530291100, Bibcode 2003PNAS..100.3983A).
- O'Brien CA, Pollett A, Gallinger S, Dick JE, « A human colon cancer cell capable of initiating tumour growth in immunodeficient mice », Nature, vol. 445, no 7123, , p. 106–10 (PMID 17122772, DOI 10.1038/nature05372, Bibcode 2007Natur.445..106O)
- Zhang S, Balch C, Chan MW, Lai HC, Matei D, Schilder JM, Yan PS, Huang TH, Nephew KP, « Identification and characterization of ovarian cancer-initiating cells from primary human tumors », Cancer Research, vol. 68, no 11, , p. 4311–20 (PMID 18519691, PMCID 2553722, DOI 10.1158/0008-5472.CAN-08-0364)
- Alvero AB, Chen R, Fu HH, Montagna M, Schwartz PE, Rutherford T, Silasi DA, Steffensen KD, Waldstrom M, Visintin I, Mor G, « Molecular phenotyping of human ovarian cancer stem cells unravels the mechanisms for repair and chemoresistance », Cell Cycle, vol. 8, no 1, , p. 158–66 (PMID 19158483, PMCID 3041590, DOI 10.4161/cc.8.1.7533)
- Li C, Heidt DG, Dalerba P, Burant CF, Zhang L, Adsay V, Wicha M, Clarke MF, Simeone DM, « Identification of pancreatic cancer stem cells », Cancer Research, vol. 67, no 3, , p. 1030–7 (PMID 17283135, DOI 10.1158/0008-5472.CAN-06-2030)
- Maitland NJ, Collins AT, « Prostate cancer stem cells: a new target for therapy », Journal of Clinical Oncology, vol. 26, no 17, , p. 2862–70 (PMID 18539965, DOI 10.1200/JCO.2007.15.1472)
- Lang SH, Frame FM, Collins AT, « Prostate cancer stem cells », The Journal of Pathology, vol. 217, no 2, , p. 299–306 (PMID 19040209, PMCID 2673349, DOI 10.1002/path.2478)
- Schatton T, Murphy GF, Frank NY, Yamaura K, Waaga-Gasser AM, Gasser M, Zhan Q, Jordan S, Duncan LM, Weishaupt C, Fuhlbrigge RC, Kupper TS, Sayegh MH, Frank MH, « Identification of cells initiating human melanomas », Nature, vol. 451, no 7176, , p. 345–9 (PMID 18202660, PMCID 3660705, DOI 10.1038/nature06489, Bibcode 2008Natur.451..345S)
- Boiko AD, Razorenova OV, van de Rijn M, Swetter SM, Johnson DL, Ly DP, Butler PD, Yang GP, Joshua B, Kaplan MJ, Longaker MT, Weissman IL, « Human melanoma-initiating cells express neural crest nerve growth factor receptor CD271 », Nature, vol. 466, no 7302, , p. 133–7 (PMID 20596026, PMCID 2898751, DOI 10.1038/nature09161, Bibcode 2010Natur.466..133B)
- Schmidt P, Kopecky C, Hombach A, Zigrino P, Mauch C, Abken H, « Eradication of melanomas by targeted elimination of a minor subset of tumor cells », Proceedings of the National Academy of Sciences of the United States of America, vol. 108, no 6, , p. 2474–9 (PMID 21282657, PMCID 3038763, DOI 10.1073/pnas.1009069108, Bibcode 2011PNAS..108.2474S, lire en ligne)
- Civenni G, Walter A, Kobert N, Mihic-Probst D, Zipser M, Belloni B, Seifert B, Moch H, Dummer R, van den Broek M, Sommer L, « Human CD271-positive melanoma stem cells associated with metastasis establish tumor heterogeneity and long-term growth », Cancer Research, vol. 71, no 8, , p. 3098–109 (PMID 21393506, DOI 10.1158/0008-5472.CAN-10-3997, lire en ligne)
- Matsui W, Huff CA, Wang Q, Malehorn MT, Barber J, Tanhehco Y, Smith BD, Civin CI, Jones RJ, « Characterization of clonogenic multiple myeloma cells », Blood, vol. 103, no 6, , p. 2332–6 (PMID 14630803, PMCID 3311914, DOI 10.1182/blood-2003-09-3064)
- Matsui W, Wang Q, Barber JP, Brennan S, Smith BD, Borrello I, McNiece I, Lin L, Ambinder RF, Peacock C, Watkins DN, Huff CA, Jones RJ, « Clonogenic multiple myeloma progenitors, stem cell properties, and drug resistance », Cancer Research, vol. 68, no 1, , p. 190–7 (PMID 18172311, PMCID 2603142, DOI 10.1158/0008-5472.CAN-07-3096)
- Colmont CS, Benketah A, Reed SH, Hawk NV, Telford WG, Ohyama M, Udey MC, Yee CL, Vogel JC, Patel GK, « CD200-expressing human basal cell carcinoma cells initiate tumor growth », Proceedings of the National Academy of Sciences of the United States of America, vol. 110, no 4, , p. 1434–9 (PMID 23292936, PMCID 3557049, DOI 10.1073/pnas.1211655110, Bibcode 2013PNAS..110.1434C)
- Patel GK, Yee CL, Terunuma A, Telford WG, Voong N, Yuspa SH, Vogel JC, « Identification and characterization of tumor-initiating cells in human primary cutaneous squamous cell carcinoma », The Journal of Investigative Dermatology, vol. 132, no 2, , p. 401–9 (PMID 22011906, PMCID 3258300, DOI 10.1038/jid.2011.317)
- Preziosi, Luigi, Cancer Modelling and Simulation, Boca Raton, CRC Press, , 456 p. (ISBN 978-1-58488-361-6)
- Ganguly R, Puri IK, « Mathematical model for the cancer stem cell hypothesis », Cell Proliferation, vol. 39, no 1, , p. 3–14 (PMID 16426418, PMCID 6495990, DOI 10.1111/j.1365-2184.2006.00369.x)
- Ganguly R, Puri IK, « Mathematical model for chemotherapeutic drug efficacy in arresting tumour growth based on the cancer stem cell hypothesis », Cell Proliferation, vol. 40, no 3, , p. 338–54 (PMID 17531079, PMCID 6496895, DOI 10.1111/j.1365-2184.2007.00434.x)
- Wang Y, Yang J, Zheng H, Tomasek GJ, Zhang P, McKeever PE, Lee EY, Zhu Y, « Expression of mutant p53 proteins implicates a lineage relationship between neural stem cells and malignant astrocytic glioma in a murine model », Cancer Cell, vol. 15, no 6, , p. 514–26 (PMID 19477430, PMCID 2721466, DOI 10.1016/j.ccr.2009.04.001)
- López-Lázaro M, « The migration ability of stem cells can explain the existence of cancer of unknown primary site. Rethinking metastasis », Oncoscience, vol. 2, no 5, , p. 467–75 (PMID 26097879, PMCID 4468332, DOI 10.18632/oncoscience.159)
- López-Lázaro M, « Stem cell division theory of cancer », Cell Cycle, vol. 14, no 16, , p. 2547–8 (PMID 26090957, PMCID 5242319, DOI 10.1080/15384101.2015.1062330)
- Clarke MF, Dick JE, Dirks PB, Eaves CJ, Jamieson CH, Jones DL, Visvader J, Weissman IL, Wahl GM, « Cancer stem cells--perspectives on current status and future directions: AACR Workshop on cancer stem cells », Cancer Research, vol. 66, no 19, , p. 9339–44 (PMID 16990346, DOI 10.1158/0008-5472.CAN-06-3126)
- Golebiewska A, Brons NH, Bjerkvig R, Niclou SP, « Critical appraisal of the side population assay in stem cell and cancer stem cell research », Cell Stem Cell, vol. 8, no 2, , p. 136–47 (PMID 21295271, DOI 10.1016/j.stem.2011.01.007)
- Scharenberg CW, Harkey MA, Torok-Storb B, « The ABCG2 transporter is an efficient Hoechst 33342 efflux pump and is preferentially expressed by immature human hematopoietic progenitors », Blood, vol. 99, no 2, , p. 507–12 (PMID 11781231, DOI 10.1182/blood.V99.2.507)
- Pastrana E, Silva-Vargas V, Doetsch F, « Eyes wide open: a critical review of sphere-formation as an assay for stem cells », Cell Stem Cell, vol. 8, no 5, , p. 486–98 (PMID 21549325, PMCID 3633588, DOI 10.1016/j.stem.2011.04.007)
- Nicolis SK, « Cancer stem cells and "stemness" genes in neuro-oncology », Neurobiology of Disease, vol. 25, no 2, , p. 217–29 (PMID 17141509, DOI 10.1016/j.nbd.2006.08.022)
- Kim YS, Kaidina AM, Chiang JH, Yarygin KN, Lupatov AY, « Cancer stem cell molecular markers verified in vivo », Biochem. Moscow Suppl. Ser. B, vol. 11, no 1, , p. 43–54 (DOI 10.1134/S1990750817010036)
- Meng E, Mitra A, Tripathi K, Finan MA, Scalici J, McClellan S, Madeira da Silva L, Reed E, Shevde LA, Palle K, Rocconi RP, « ALDH1A1 maintains ovarian cancer stem cell-like properties by altered regulation of cell cycle checkpoint and DNA repair network signaling », PLOS ONE, vol. 9, no 9, , e107142 (PMID 25216266, PMCID 4162571, DOI 10.1371/journal.pone.0107142)
- Visvader JE, Lindeman GJ, « Cancer stem cells in solid tumours: accumulating evidence and unresolved questions », Nature Reviews. Cancer, vol. 8, no 10, , p. 755–68 (PMID 18784658, DOI 10.1038/nrc2499)
- Hirschmann-Jax C, Foster AE, Wulf GG, Nuchtern JG, Jax TW, Gobel U, Goodell MA, Brenner MK, « A distinct "side population" of cells with high drug efflux capacity in human tumor cells », Proceedings of the National Academy of Sciences of the United States of America, vol. 101, no 39, , p. 14228–33 (PMID 15381773, PMCID 521140, DOI 10.1073/pnas.0400067101, Bibcode 2004PNAS..10114228H)
- Ginestier C, Hur MH, Charafe-Jauffret E, Monville F, Dutcher J, Brown M, Jacquemier J, Viens P, Kleer CG, Liu S, Schott A, Hayes D, Birnbaum D, Wicha MS, Dontu G, « ALDH1 is a marker of normal and malignant human mammary stem cells and a predictor of poor clinical outcome », Cell Stem Cell, vol. 1, no 5, , p. 555–67 (PMID 18371393, PMCID 2423808, DOI 10.1016/j.stem.2007.08.014)
- Pece S, Tosoni D, Confalonieri S, Mazzarol G, Vecchi M, Ronzoni S, Bernard L, Viale G, Pelicci PG, Di Fiore PP, « Biological and molecular heterogeneity of breast cancers correlates with their cancer stem cell content », Cell, vol. 140, no 1, , p. 62–73 (PMID 20074520, DOI 10.1016/j.cell.2009.12.007)
- Deng S, Yang X, Lassus H, Liang S, Kaur S, Ye Q, Li C, Wang LP, Roby KF, Orsulic S, Connolly DC, Zhang Y, Montone K, Bützow R, Coukos G, Zhang L, « Distinct expression levels and patterns of stem cell marker, aldehyde dehydrogenase isoform 1 (ALDH1), in human epithelial cancers », PLOS ONE, vol. 5, no 4, , e10277 (PMID 20422001, PMCID 2858084, DOI 10.1371/journal.pone.0010277, Bibcode 2010PLoSO...510277D)
- Meyer MJ, Fleming JM, Lin AF, Hussnain SA, Ginsburg E, Vonderhaar BK, « CD44posCD49fhiCD133/2hi defines xenograft-initiating cells in estrogen receptor-negative breast cancer », Cancer Research, vol. 70, no 11, , p. 4624–33 (PMID 20484027, PMCID 4129519, DOI 10.1158/0008-5472.CAN-09-3619)
- Singh SK, Hawkins C, Clarke ID, Squire JA, Bayani J, Hide T, Henkelman RM, Cusimano MD, Dirks PB, « Identification of human brain tumour initiating cells », Nature, vol. 432, no 7015, , p. 396–401 (PMID 15549107, DOI 10.1038/nature03128, Bibcode 2004Natur.432..396S)
- Son MJ, Woolard K, Nam DH, Lee J, Fine HA, « SSEA-1 is an enrichment marker for tumor-initiating cells in human glioblastoma », Cell Stem Cell, vol. 4, no 5, , p. 440–52 (PMID 19427293, PMCID 7227614, DOI 10.1016/j.stem.2009.03.003)
- Mazzoleni S, Politi LS, Pala M, Cominelli M, Franzin A, Sergi Sergi L, Falini A, De Palma M, Bulfone A, Poliani PL, Galli R, « Epidermal growth factor receptor expression identifies functionally and molecularly distinct tumor-initiating cells in human glioblastoma multiforme and is required for gliomagenesis », Cancer Research, vol. 70, no 19, , p. 7500–13 (PMID 20858720, DOI 10.1158/0008-5472.CAN-10-2353, lire en ligne)
- Anido J, Sáez-Borderías A, Gonzàlez-Juncà A, Rodón L, Folch G, Carmona MA, Prieto-Sánchez RM, Barba I, Martínez-Sáez E, Prudkin L, Cuartas I, Raventós C, Martínez-Ricarte F, Poca MA, García-Dorado D, Lahn MM, Yingling JM, Rodón J, Sahuquillo J, Baselga J, Seoane J, « TGF-β Receptor Inhibitors Target the CD44(high)/Id1(high) Glioma-Initiating Cell Population in Human Glioblastoma », Cancer Cell, vol. 18, no 6, , p. 655–68 (PMID 21156287, DOI 10.1016/j.ccr.2010.10.023)
- Dalerba P, Dylla SJ, Park IK, Liu R, Wang X, Cho RW, Hoey T, Gurney A, Huang EH, Simeone DM, Shelton AA, Parmiani G, Castelli C, Clarke MF, « Phenotypic characterization of human colorectal cancer stem cells », Proceedings of the National Academy of Sciences of the United States of America, vol. 104, no 24, , p. 10158–63 (PMID 17548814, PMCID 1891215, DOI 10.1073/pnas.0703478104, Bibcode 2007PNAS..10410158D)
- Wilson BJ, Schatton T, Zhan Q, Gasser M, Ma J, Saab KR, Schanche R, Waaga-Gasser AM, Gold JS, Huang Q, Murphy GF, Frank MH, Frank NY, « ABCB5 identifies a therapy-refractory tumor cell population in colorectal cancer patients », Cancer Research, vol. 71, no 15, , p. 5307–16 (PMID 21652540, PMCID 3395026, DOI 10.1158/0008-5472.CAN-11-0221)
- Odoux C, Fohrer H, Hoppo T, Guzik L, Stolz DB, Lewis DW, Gollin SM, Gamblin TC, Geller DA, Lagasse E, « A stochastic model for cancer stem cell origin in metastatic colon cancer », Cancer Research, vol. 68, no 17, , p. 6932–41 (PMID 18757407, PMCID 2562348, DOI 10.1158/0008-5472.CAN-07-5779)
- Huang EH, Hynes MJ, Zhang T, Ginestier C, Dontu G, Appelman H, Fields JZ, Wicha MS, Boman BM, « Aldehyde dehydrogenase 1 is a marker for normal and malignant human colonic stem cells (SC) and tracks SC overpopulation during colon tumorigenesis », Cancer Research, vol. 69, no 8, , p. 3382–9 (PMID 19336570, PMCID 2789401, DOI 10.1158/0008-5472.CAN-08-4418)
- Kemper K, Sprick MR, de Bree M, Scopelliti A, Vermeulen L, Hoek M, Zeilstra J, Pals ST, Mehmet H, Stassi G, Medema JP, « The AC133 epitope, but not the CD133 protein, is lost upon cancer stem cell differentiation », Cancer Research, vol. 70, no 2, , p. 719–29 (PMID 20068153, DOI 10.1158/0008-5472.CAN-09-1820)
- Liu C, Kelnar K, Liu B, Chen X, Calhoun-Davis T, Li H, Patrawala L, Yan H, Jeter C, Honorio S, Wiggins JF, Bader AG, Fagin R, Brown D, Tang DG, « The microRNA miR-34a inhibits prostate cancer stem cells and metastasis by directly repressing CD44 », Nature Medicine, vol. 17, no 2, , p. 211–5 (PMID 21240262, PMCID 3076220, DOI 10.1038/nm.2284)
- Ho MM, Ng AV, Lam S, Hung JY, « Side population in human lung cancer cell lines and tumors is enriched with stem-like cancer cells », Cancer Research, vol. 67, no 10, , p. 4827–33 (PMID 17510412, DOI 10.1158/0008-5472.CAN-06-3557)
- Patrawala L, Calhoun T, Schneider-Broussard R, Li H, Bhatia B, Tang S, Reilly JG, Chandra D, Zhou J, Claypool K, Coghlan L, Tang DG, « Highly purified CD44+ prostate cancer cells from xenograft human tumors are enriched in tumorigenic and metastatic progenitor cells », Oncogene, vol. 25, no 12, , p. 1696–708 (PMID 16449977, DOI 10.1038/sj.onc.1209327)
- Dubrovska A, Kim S, Salamone RJ, Walker JR, Maira SM, García-Echeverría C, Schultz PG, Reddy VA, « The role of PTEN/Akt/PI3K signaling in the maintenance and viability of prostate cancer stem-like cell populations », Proceedings of the National Academy of Sciences of the United States of America, vol. 106, no 1, , p. 268–73 (PMID 19116269, PMCID 2629188, DOI 10.1073/pnas.0810956106, Bibcode 2009PNAS..106..268D)
- Rajasekhar VK, Studer L, Gerald W, Socci ND, Scher HI, « Tumour-initiating stem-like cells in human prostate cancer exhibit increased NF-κB signalling », Nature Communications, vol. 2, no 1, , p. 162 (PMID 21245843, PMCID 3105310, DOI 10.1038/ncomms1159, Bibcode 2011NatCo...2..162R)
- Li T, Su Y, Mei Y, Leng Q, Leng B, Liu Z, Stass SA, Jiang F, « ALDH1A1 is a marker for malignant prostate stem cells and predictor of prostate cancer patients' outcome », Laboratory Investigation; A Journal of Technical Methods and Pathology, vol. 90, no 2, , p. 234–44 (PMID 20010854, PMCID 3552330, DOI 10.1038/labinvest.2009.127)
- Eramo A, Lotti F, Sette G, Pilozzi E, Biffoni M, Di Virgilio A, Conticello C, Ruco L, Peschle C, De Maria R, « Identification and expansion of the tumorigenic lung cancer stem cell population », Cell Death and Differentiation, vol. 15, no 3, , p. 504–14 (PMID 18049477, DOI 10.1038/sj.cdd.4402283)
- Sullivan JP, Spinola M, Dodge M, Raso MG, Behrens C, Gao B, Schuster K, Shao C, Larsen JE, Sullivan LA, Honorio S, Xie Y, Scaglioni PP, DiMaio JM, Gazdar AF, Shay JW, Wistuba II, Minna JD, « Aldehyde dehydrogenase activity selects for lung adenocarcinoma stem cells dependent on notch signaling », Cancer Research, vol. 70, no 23, , p. 9937–48 (PMID 21118965, PMCID 3058307, DOI 10.1158/0008-5472.CAN-10-0881)
- Leung EL, Fiscus RR, Tung JW, Tin VP, Cheng LC, Sihoe AD, Fink LM, Ma Y, Wong MP, « Non-small cell lung cancer cells expressing CD44 are enriched for stem cell-like properties », PLOS ONE, vol. 5, no 11, , e14062 (PMID 21124918, PMCID 2988826, DOI 10.1371/journal.pone.0014062, Bibcode 2010PLoSO...514062L)
- Damelin M, Geles KG, Follettie MT, Yuan P, Baxter M, Golas J, DiJoseph JF, Karnoub M, Huang S, Diesl V, Behrens C, Choe SE, Rios C, Gruzas J, Sridharan L, Dougher M, Kunz A, Hamann PR, Evans D, Armellino D, Khandke K, Marquette K, Tchistiakova L, Boghaert ER, Abraham RT, Wistuba II, Zhou BB, « Delineation of a cellular hierarchy in lung cancer reveals an oncofetal antigen expressed on tumor-initiating cells », Cancer Research, vol. 71, no 12, , p. 4236–46 (PMID 21540235, DOI 10.1158/0008-5472.CAN-10-3919)
- Thiery JP, « Epithelial-mesenchymal transitions in tumour progression », Nature Reviews. Cancer, vol. 2, no 6, , p. 442–54 (PMID 12189386, DOI 10.1038/nrc822)
- Angerer LM, Angerer RC, « Regulative development of the sea urchin embryo: signalling cascades and morphogen gradients », Seminars in Cell & Developmental Biology, vol. 10, no 3, , p. 327–34 (PMID 10441547, DOI 10.1006/scdb.1999.0292)
- Mani SA, Yang J, Brooks M, Schwaninger G, Zhou A, Miura N, Kutok JL, Hartwell K, Richardson AL, Weinberg RA, « Mesenchyme Forkhead 1 (FOXC2) plays a key role in metastasis and is associated with aggressive basal-like breast cancers », Proceedings of the National Academy of Sciences of the United States of America, vol. 104, no 24, , p. 10069–74 (PMID 17537911, PMCID 1891217, DOI 10.1073/pnas.0703900104, Bibcode 2007PNAS..10410069M)
- Conacci-Sorrell M, Simcha I, Ben-Yedidia T, Blechman J, Savagner P, Ben-Ze'ev A, « Autoregulation of E-cadherin expression by cadherin-cadherin interactions: the roles of beta-catenin signaling, Slug, and MAPK », The Journal of Cell Biology, vol. 163, no 4, , p. 847–57 (PMID 14623871, PMCID 2173691, DOI 10.1083/jcb.200308162)
- Comito G, Calvani M, Giannoni E, Bianchini F, Calorini L, Torre E, Migliore C, Giordano S, Chiarugi P, « HIF-1α stabilization by mitochondrial ROS promotes Met-dependent invasive growth and vasculogenic mimicry in melanoma cells », Free Radical Biology & Medicine, vol. 51, no 4, , p. 893–904 (PMID 21703345, DOI 10.1016/j.freeradbiomed.2011.05.042, hdl 2158/496457)
- Spill F, Reynolds DS, Kamm RD, Zaman MH, « Impact of the physical microenvironment on tumor progression and metastasis », Current Opinion in Biotechnology, vol. 40, , p. 41–48 (PMID 26938687, PMCID 4975620, DOI 10.1016/j.copbio.2016.02.007)
- Kaplan RN, Riba RD, Zacharoulis S, Bramley AH, Vincent L, Costa C, MacDonald DD, Jin DK, Shido K, Kerns SA, Zhu Z, Hicklin D, Wu Y, Port JL, Altorki N, Port ER, Ruggero D, Shmelkov SV, Jensen KK, Rafii S, Lyden D, « VEGFR1-positive haematopoietic bone marrow progenitors initiate the pre-metastatic niche », Nature, vol. 438, no 7069, , p. 820–7 (PMID 16341007, PMCID 2945882, DOI 10.1038/nature04186, Bibcode 2005Natur.438..820K)
- Hermann PC, Huber SL, Herrler T, Aicher A, Ellwart JW, Guba M, Bruns CJ, Heeschen C, « Distinct populations of cancer stem cells determine tumor growth and metastatic activity in human pancreatic cancer », Cell Stem Cell, vol. 1, no 3, , p. 313–23 (PMID 18371365, DOI 10.1016/j.stem.2007.06.002)
- Yang ZF, Ho DW, Ng MN, Lau CK, Yu WC, Ngai P, Chu PW, Lam CT, Poon RT, Fan ST, « Significance of CD90+ cancer stem cells in human liver cancer », Cancer Cell, vol. 13, no 2, , p. 153–66 (PMID 18242515, DOI 10.1016/j.ccr.2008.01.013)
- Shipitsin M, Campbell LL, Argani P, Weremowicz S, Bloushtain-Qimron N, Yao J, Nikolskaya T, Serebryiskaya T, Beroukhim R, Hu M, Halushka MK, Sukumar S, Parker LM, Anderson KS, Harris LN, Garber JE, Richardson AL, Schnitt SJ, Nikolsky Y, Gelman RS, Polyak K, « Molecular definition of breast tumor heterogeneity », Cancer Cell, vol. 11, no 3, , p. 259–73 (PMID 17349583, DOI 10.1016/j.ccr.2007.01.013)
- Shmelkov SV, Butler JM, Hooper AT, Hormigo A, Kushner J, Milde T, St Clair R, Baljevic M, White I, Jin DK, Chadburn A, Murphy AJ, Valenzuela DM, Gale NW, Thurston G, Yancopoulos GD, D'Angelica M, Kemeny N, Lyden D, Rafii S, « CD133 expression is not restricted to stem cells, and both CD133+ and CD133- metastatic colon cancer cells initiate tumors », The Journal of Clinical Investigation, vol. 118, no 6, , p. 2111–20 (PMID 18497886, PMCID 2391278, DOI 10.1172/JCI34401)
- Brabletz T, Jung A, Spaderna S, Hlubek F, Kirchner T, « Opinion: migrating cancer stem cells - an integrated concept of malignant tumour progression », Nature Reviews. Cancer, vol. 5, no 9, , p. 744–9 (PMID 16148886, DOI 10.1038/nrc1694)
- Zhao J, « Cancer stem cells and chemoresistance: The smartest survives the raid », Pharmacol Ther, vol. 160, , p. 145–58 (PMID 26899500, PMCID 4808328, DOI 10.1016/j.pharmthera.2016.02.008)
- Cyril Corbet et Alexandre Prieur, Therapeutic Targeting of Cancer Stem-Like Cells (CSC) – The Current State of the Art, Frontiers Media SA, (ISBN 978-2-88963-647-1, lire en ligne)
- Dirks P, « Cancer stem cells: Invitation to a second round », Nature, vol. 466, no 7302, , p. 40–1 (PMID 20596007, DOI 10.1038/466040a, Bibcode 2010Natur.466...40D)
- Haskins WE, Eedala S, Jadhav YL, Labhan MS, Pericherla VC, Perlman EJ, « Insights on neoplastic stem cells from gel-based proteomics of childhood germ cell tumors », Pediatric Blood & Cancer, vol. 58, no 5, , p. 722–8 (PMID 21793190, PMCID 3204330, DOI 10.1002/pbc.23282)
- Gupta PB, Onder TT, Jiang G, Tao K, Kuperwasser C, Weinberg RA, Lander ES, « Identification of selective inhibitors of cancer stem cells by high-throughput screening », Cell, vol. 138, no 4, , p. 645–659 (PMID 19682730, PMCID 4892125, DOI 10.1016/j.cell.2009.06.034)
- Jangamreddy JR, Ghavami S, Grabarek J, Kratz G, Wiechec E, Fredriksson BA, Rao Pariti RK, Cieślar-Pobuda A, Panigrahi S, Łos MJ, « Salinomycin induces activation of autophagy, mitophagy and affects mitochondrial polarity: differences between primary and cancer cells », Biochimica et Biophysica Acta (BBA) - Molecular Cell Research, vol. 1833, no 9, , p. 2057–69 (PMID 23639289, DOI 10.1016/j.bbamcr.2013.04.011)
- Vlahopoulos S, Critselis E, Voutsas IF, Perez SA, Moschovi M, Baxevanis CN, Chrousos GP, « New use for old drugs? Prospective targets of chloroquines in cancer therapy », Current Drug Targets, vol. 15, no 9, , p. 843–51 (PMID 25023646, DOI 10.2174/1389450115666140714121514)
- Jordan CT, Upchurch D, Szilvassy SJ, Guzman ML, Howard DS, Pettigrew AL, Meyerrose T, Rossi R, Grimes B, Rizzieri DA, Luger SM, Phillips GL, « The interleukin-3 receptor alpha chain is a unique marker for human acute myelogenous leukemia stem cells », Leukemia, vol. 14, no 10, , p. 1777–84 (PMID 11021753, DOI 10.1038/sj.leu.2401903)
- Jin L, Lee EM, Ramshaw HS, Busfield SJ, Peoppl AG, Wilkinson L, Guthridge MA, Thomas D, Barry EF, Boyd A, Gearing DP, Vairo G, Lopez AF, Dick JE, Lock RB, « Monoclonal antibody-mediated targeting of CD123, IL-3 receptor alpha chain, eliminates human acute myeloid leukemic stem cells », Cell Stem Cell, vol. 5, no 1, , p. 31–42 (PMID 19570512, DOI 10.1016/j.stem.2009.04.018)
- « 'Nanobombs' that blow up cancer cells | KurzweilAI », sur www.kurzweilai.net, (consulté le )
- Wang H, Agarwal P, Zhao S, Yu J, Lu X, He X, « A Near-Infrared Laser-Activated "Nanobomb" for Breaking the Barriers to MicroRNA Delivery », Advanced Materials, vol. 28, no 2, , p. 347–55 (PMID 26567892, DOI 10.1002/adma.201504263)
- Huddle BC, Grimley E, Buchman CD, Chtcherbinine M, Debnath B, Mehta P, Yang K, Morgan CA, Li S, Felton JA, Sun D, Metha G, Neamati N, Buckanovich RJ, Hurley TD, Larsen SD, « Structure-Based Optimization of a Novel Class of Aldehyde Dehydrogenase 1A (ALDH1A) Subfamily-Selective Inhibitors as Potential Adjuncts to Ovarian Cancer Chemotherapy », J Med Chem, vol. 61, no 19, , p. 8754–8773 (PMID 30221940, PMCID 6477540, DOI 10.1021/acs.jmedchem.8b00930)
- Haupt Y, Bath ML, Harris AW, Adams JM, « bmi-1 transgene induces lymphomas and collaborates with myc in tumorigenesis », Oncogene, vol. 8, no 11, , p. 3161–4 (PMID 8414519)
- Park IK, Qian D, Kiel M, Becker MW, Pihalja M, Weissman IL, Morrison SJ, Clarke MF, « Bmi-1 is required for maintenance of adult self-renewing haematopoietic stem cells », Nature, vol. 423, no 6937, , p. 302–5 (PMID 12714971, DOI 10.1038/nature01587, Bibcode 2003Natur.423..302P, hdl 2027.42/62508, lire en ligne)
- Molofsky AV, Pardal R, Iwashita T, Park IK, Clarke MF, Morrison SJ, « Bmi-1 dependence distinguishes neural stem cell self-renewal from progenitor proliferation », Nature, vol. 425, no 6961, , p. 962–7 (PMID 14574365, PMCID 2614897, DOI 10.1038/nature02060, Bibcode 2003Natur.425..962M)
- Hemmati HD, Nakano I, Lazareff JA, Masterman-Smith M, Geschwind DH, Bronner-Fraser M, Kornblum HI, « Cancerous stem cells can arise from pediatric brain tumors », Proceedings of the National Academy of Sciences of the United States of America, vol. 100, no 25, , p. 15178–83 (PMID 14645703, PMCID 299944, DOI 10.1073/pnas.2036535100, Bibcode 2003PNAS..10015178H)
- Dontu G, Jackson KW, McNicholas E, Kawamura MJ, Abdallah WM, Wicha MS, « Role of Notch signaling in cell-fate determination of human mammary stem/progenitor cells », Breast Cancer Research, vol. 6, no 6, , R605–15 (PMID 15535842, PMCID 1064073, DOI 10.1186/bcr920)
- Diévart A, Beaulieu N, Jolicoeur P, « Involvement of Notch1 in the development of mouse mammary tumors », Oncogene, vol. 18, no 44, , p. 5973–81 (PMID 10557086, DOI 10.1038/sj.onc.1202991)
- Park DM, Jung J, Masjkur J, Makrogkikas S, Ebermann D, Saha S, Rogliano R, Paolillo N, Pacioni S, McKay RD, Poser S, Androutsellis-Theotokis A, « Hes3 regulates cell number in cultures from glioblastoma multiforme with stem cell characteristics », Scientific Reports, vol. 3, , p. 1095 (PMID 23393614, PMCID 3566603, DOI 10.1038/srep01095, Bibcode 2013NatSR...3E1095P)
- Beachy PA, Karhadkar SS, Berman DM, « Tissue repair and stem cell renewal in carcinogenesis », Nature, vol. 432, no 7015, , p. 324–31 (PMID 15549094, DOI 10.1038/nature03100, Bibcode 2004Natur.432..324B)
- Zhou BP, Hung MC, « Wnt, hedgehog and snail: sister pathways that control by GSK-3beta and beta-Trcp in the regulation of metastasis », Cell Cycle, vol. 4, no 6, , p. 772–6 (PMID 15917668, DOI 10.4161/cc.4.6.1744, lire en ligne)
- Akiyoshi T, Nakamura M, Koga K, Nakashima H, Yao T, Tsuneyoshi M, Tanaka M, Katano M, « Gli1, downregulated in colorectal cancers, inhibits proliferation of colon cancer cells involving Wnt signalling activation », Gut, vol. 55, no 7, , p. 991–9 (PMID 16299030, PMCID 1856354, DOI 10.1136/gut.2005.080333)
- She M, Chen X, « Targeting Signal Pathways active in Cancer Stem Cells to Overcome Drug Resistance », Zhongguo Fei AI Za Zhi = Chinese Journal of Lung Cancer, vol. 12, no 1, , p. 3–7 (PMID 20712949, DOI 10.3779/j.issn.1009-3419.2009.01.001)
- Bollmann FM, « The many faces of telomerase: emerging extratelomeric effects », BioEssays, vol. 30, no 8, , p. 728–32 (PMID 18623070, DOI 10.1002/bies.20793)
- Russ Hodge, « Hacking the programs of cancer stem cells », sur medicalxpress.com, Medical Express, (consulté le )
- Masuko Katoh et Masaru Katoh, « Molecular genetics and targeted therapy of WNT-related human diseases (Review) », International Journal of Molecular Medicine, vol. 40, no 3, , p. 587–606 (ISSN 1107-3756, PMID 28731148, PMCID PMC5547940, DOI 10.3892/ijmm.2017.3071, lire en ligne, consulté le )
- (en) Frédéric Hollande, Debra J. Lee, Armelle Choquet et Serge Roche, « Adherens junctions and tight junctions are regulated via different pathways by progastrin in epithelial cells », Journal of Cell Science, vol. 116, no 7, , p. 1187–1197 (ISSN 0021-9533 et 1477-9137, PMID 12615962, DOI 10.1242/jcs.00321, lire en ligne, consulté le )
- (en) Singh P et Owlia A, « Gastrin Gene Expression Is Required for the Proliferation and Tumorigenicity of Human Colon Cancer Cells », sur Cancer research, (PMID 8797575, consulté le )
- Hai Wu, Azarmidokht Owlia et Pomila Singh, « Precursor peptide progastrin1-80 reduces apoptosis of intestinal epithelial cells and upregulates cytochrome c oxidase Vb levels and synthesis of ATP », American Journal of Physiology-Gastrointestinal and Liver Physiology, vol. 285, no 6, , G1097–G1110 (ISSN 0193-1857, DOI 10.1152/ajpgi.00216.2003, lire en ligne, consulté le )
- (en) Alexandre Prieur, Monica Cappellini, Guillaume Habif, Marie-Paule Lefranc, Thibault Mazard, Eric Morency, Jean-Marc Pascussi, Nathalie Cahuzac, Berengere Vire, Benjamin Dubuc, Amandine Durochat, Pierre Liaud, Jeremy Ollier, Caroline Pfeiffer, Sophie Poupeau, Veronique Saywell, Chris Planque, Eric Assenat, Frederic Bibeau, Jean-François Bourgaux, Pascal Pujol, Alain Sezeur, Marc Ychou et Dominique Joubert, « Targeting the Wnt Pathway and Cancer Stem Cells with Anti-progastrin Humanized Antibodies as a Potential Treatment for K-RAS-Mutated Colorectal Cancer », Clinical Cancer Research, vol. 23, no 17, , p. 5267–5280 (ISSN 1078-0432 et 1557-3265, PMID 28600477, DOI 10.1158/1078-0432.CCR-17-0533, lire en ligne, consulté le )
- (en) Julie Giraud, Laura M. Failla, Jean-Marc Pascussi, Ebba L. Lagerqvist, Jérémy Ollier, Pascal Finetti, François Bertucci, Chu Ya, Imène Gasmi, Jean-François Bourgaux, Michel Prudhomme, Thibault Mazard, Imade Ait-Arsa, Leila Houhou, Daniel Birnbaum, André Pélegrin, Charles Vincent, James G Ryall, Dominique Joubert, Julie Pannequin et Frédéric Hollande, « Autocrine Secretion of Progastrin Promotes the Survival and Self-Renewal of Colon Cancer Stem–like Cells », Cancer Research, vol. 76, no 12, , p. 3618–3628 (ISSN 0008-5472 et 1538-7445, PMID 27197176, DOI 10.1158/0008-5472.CAN-15-1497, lire en ligne, consulté le )
- (en) S Najib, A Kowalski-Chauvel, C Do et S Roche, « Progastrin a new pro-angiogenic factor in colorectal cancer », Oncogene, vol. 34, no 24, , p. 3120–3130 (ISSN 0950-9232 et 1476-5594, DOI 10.1038/onc.2014.255, lire en ligne, consulté le )
- Benoit You, Frédéric Mercier, Eric Assenat, Carole Langlois-Jacques, Olivier Glehen, Julien Soulé, Léa Payen, Vahan Kepenekian, Marie Dupuy, anny Belouin, Eric Morency, Véronique Saywell, Maud Flacelière, Philippe Elies, Pierre Liaud, Thibault Mazard, Delphine Maucort-Boulch, Winston Tan, Bérengère Vire, Laurent Villeneuve, Marc Ychou, Manish Kohli, Dominique Joubert et Alexandre Prieur, « The oncogenic and druggable hPG80 (Progastrin) is overexpressed in multiple cancers and detected in the blood of patients », EBioMedicine, vol. 51, , p. 102574 (ISSN 2352-3964, PMID 31877416, PMCID PMC6938867, DOI 10.1016/j.ebiom.2019.11.035, lire en ligne, consulté le )
Lectures complémentaires
- Rajasekhar Vinagolu K., Cancer Stem Cells, Hoboken, New Jersey, Wiley, , 552 p. (ISBN 978-1-118-35616-6)
- Cyril Corbet et Alexandre Prieur, Therapeutic Targeting of Cancer Stem-Like Cells (CSC) – The Current State of the Art, Frontiers Media SA, (ISBN 978-2-88963-647-1, lire en ligne)