Campylocephalus
Campylocephalus est un genre éteint et fossile d'Euryptérides, un groupe d'Arthropodes aquatiques éteints.

| Règne | Animalia |
|---|---|
| Embranchement | Arthropoda |
| Sous-embr. | Chelicerata |
| Ordre | † Eurypterida |
| Sous-ordre | † Stylonurina |
| Super-famille | † Mycteropoidea |
| Famille | † Hibbertopteridae |
Espèces de rang inférieur
- † Campylocephalys oculatus (Kutorga, 1838)
- † Campylocephalus permianus (Ponomarenko, 1985)
- † Campylocephalus salmi (Štúr, 1877)
Présentation
Des fossiles de Campylocephalus ont été découverts dans des gisements allant du Carbonifère en République tchèque (l'espèce C. salmi) au Permien en Russie (espèces C. oculatus et C. permianus). Le nom de genre est composé des mots grecs καμπύλος (kampýlos), signifiant « courbé », et κεφαλή (kephalē), signifiant « tête ».
Campylocephalus est un membre de la famille des Euryptérides Hibbertoptérides et ressemblait probablement beaucoup aux autres membres de cette famille, Hibbertopterus et Vernonopterus, au sens où c'était un grand animal, large et et massif, assez différent des célèbres Euryptérides nageurs (tels que Pterygotus et Eurypterus) fréquents au cours des périodes antérieures. Comme tous les autres Euryptérides Stylonurines, Campylocephalus était totalement dépourvu de palettes natatoires.
Les Hibbertoptérides tels que Campylocephalus se nourrissaient, comme beaucoup d'autres familles du sous-ordre des Stylonurines, par une méthode dite d'alimentation par balayage. Les stratégies d'alimentation par balayage impliquent des appendices spécialisés dotés de lames probablement utilisées par les animaux pour ratisser le substrat de leur environnement à la recherche de petites proies.
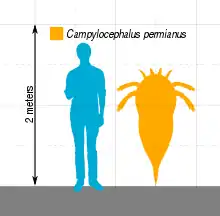
L'espèce C. permianus, découverte en Russie dans des dépôts du Permien supérieur, est la seule espèce de Campylocephalus suffisamment bien conservée pour permettre une estimation de sa taille, que plusieurs publications estiment à 1,4 mètres de long. C'est l'une des dernières espèces d'Euryptérides vivantes connues : elle s'est éteinte juste avant ou pendant l'extinction du Permien-Trias, il y a 251,9 millions d'années. Avant cette extinction massive, les Euryptérides ont connu un déclin progressif durant des millions d'années, tant en nombres et qu'en diversité : ainsi, Campylocephalus est le seul genre connu d'Euryptérides à avoir vécu au cours des plus de 20 millions d'années qui ont suivi l'extinction du genre apparenté Hastimima.
Description
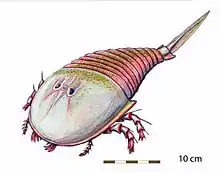
Classé dans la famille des Euryptérides Hibbertoptérides, Campylocephalus ressemble beaucoup aux autres membres de la famille. C'est une grande créature, large et massive, ce en quoi elle diffère sensiblement des plus anciens et plus célèbres Euryptérides nageurs que sont Pterygotus et Eurypterus. Comme tous les membres du sous-ordre des Stylonurines, Campylocephalus est totalement dépourvue de palettes natatoires[1] - [2].
Plusieurs traits distinguent Campylocephalus des autres genres de sa famille, en particulier du très proche Hibbertopterus. Campylocephalus a un prosome (tête) subelliptique (presque elliptique), presque semi-circulaire, fortement convexe et plus large dans sa section médiane. Les yeux composés de Campylocephalus sont positionnés latéralement (sur les côtés de la tête) et sont séparés les uns des autres par des lobes gonflés en leur milieu. Derrière les yeux, sur la carapace (la partie de l''exosquelette recouvrant la tête), on distingue d'autres structures semblables à des lobes appelées lobes palpébraux[1] - [2]
Comme le prosome, les tergites (segments visibles sur la face supérieure du corps) de l'abdomen sont de forme convexe et possèdent des processus articulaires (structures en saillie qui permettent aux segments de s'emboîter). Les appendices (membres) de Campylocephalus, très rarement conservés, sont presque totalement inconnus[1] - [2]
Compte tenu du caractère incomplet des fossiles de Campylocephalus, il est difficile de déterminer sa taille. Aucune estimation n'a été publiée concernant l'espèce type C. oculatus et l'espèce C. salmi. Le seul fossile connu de C. permianus, une carapace massive incomplète, suggère un très grand Euryptéride, qui pourrait atteindre 1,4 mètre de longueur[3].
Chez C. oculatus, les yeux de forme semi-lunaire (presque en forme de lune) sont placés près du milieu de la carapace, avec de petits ocelles (yeux simples sensibles à la lumière) entre eux. La carapace comporte de petites proéminences irrégulières, un bord antérieur arrondi et un bord postérieur échancré. Les segments thoraciques (segments du thorax ) de l'espèce sont droits et étroits[4]. Les yeux de C. salmi sont similaires à ceux de C. occulatus mais sont beaucoup plus proches l'un de l'autre[5]. Les yeux sont également un trait distinctif pour C. permianus, chez qui ils sont placés plus en arrière que chez les autres espèces de Campylocephalus[1].
Histoire de la recherche
Les fossiles désormais attribués au genre Campylocephalus ont été décrits pour la première fois en 1838 par le paléontologue russe Stepan Kutorga, qui les a alors attribués à espèce du genre Limulus, le même genre que celui de la limule moderne. En s'appuyant sur certaines similitudes anatomiques et d'aspect avec les membres du genre moderne qu'il distingue dans un spécimen fossile pourtant incomplet, Kutorga le nomme Limulus oculatus[6].
En 1831, le naturaliste écossais John Scouler décrit le genre Eidothea sur la base d'un unique prosome fossile découvert en Ecosse, sans pour autant lui attribuer de nom d'espèce. Créer ainsi un genre sans espèce va à l'encontre de la nomenclature zoologique orthodoxe, et est en contradiction avec l'Avis 65 de la Commission Internationale de Nomenclature Zoologique[7] :
« Si un auteur désigne une espèce comme espèce type d'un genre, on suppose qu'il a correctement décrit l'espèce ; si un cas se présente dans lequel il apparaît qu'un auteur a basé un genre sur certains spécimens définis plutôt que sur une espèce, il serait bon de soumettre le cas, avec tous les détails, à la Commission. Présentement, il est difficile de tirer une règle générale. »
— Avis et déclarations de la Commission Internationale de Nomenclature Zoologique (1958)[8]
Les problèmes taxonomiques nés de cette décision de Scouler ont cependant rapidement trouvé leur terme avec la prise de conscience que le nom de genre Eidothea était déjà utilisé par un genre de mollusques décrit en 1826. Néanmoins, le nom Eidothea sera également associé à l'espèce d'Euryptéride écossaise Eurypterus scouleri décrite en 1836 et qui possède un prosome presque identique à celui attribué initialement à Eidothea[7].
En 1860, le paléontologue français Edouard d'Eichwald constate que la carapace et les segments thoraciques décrits par Kutorga sont tout à fait distincts de Limulus, et donne le nom de Campylocephalus au nouveau genre destiné à accueillir Limulus oculatus. Ce nom des genre est composé des mots grecs καμπύλος (kampýlos), signifiant « courbé », et κεφαλή (kephalē), signifiant « tête »[4].
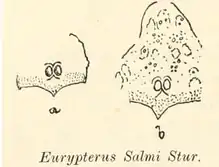
La seconde espèce de Campylocephalus décrite, en 1877, C. salmi, est initialement attribuée au genre Eurypterus, apparenté mais cependant assez éloigné, par le géologue et paléontologue slovaque Dionýs Štúr. La description de Štúr est basée sur deux carapaces fossiles incomplètes. La première carapace fragmentaire ne comporte que la partie inférieure de la tête, terminée par deux arcs brisés et concaves. Les yeux de cette carapace sont proches l'un de l'autre, positionnés au milieu de la carapace sur une partie surélevée de forme triangulaire semblable à celle de certains spécimens de l'espèce apparentée Hibbertopterus scouleri. La deuxième carapace fossile présente la particularité de commencer à s'étendre vers l'avant au-dessus des yeux. Ces deux fossiles présentent également des protubérances de différentes tailles, réparties sur l'exosquelette, de formes vagues et encore jamais vues sur les autres spécimens du genre[5].
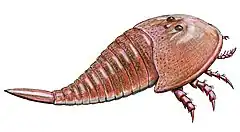
En 1951, le paléontologue norvégien Leif Størmer compare les fossiles d'Eurypterus scouleri à la carapace décrite par Kutorga en 1838 et conclut qu'ils appartiennent tous deux au même genre[9]. A cette époque, Eichwald a déjà attribué au genre Campylocephalus les fossiles initialement attribués au genre Eidothea par Scouler[7]. E. scouleri est alors renommé Campylocephalus scouleri. À cette époque, les pattes de Campylocephalus sont inconnues, malgré l'ajout de l'espèce écossaise, et tout regroupement avec d'autres genres ne peut se faire qu'en se basant sur les caractéristiques de la carapace. En comparant l'ornementation de la carapace à d'autres Euryptérides, Størmer découvre qu'elle est similaire au genre Tarsopterella (où les pattes étaient également plus ou moins inconnues) ce qui permet d'attribuer clairement Campylocephalus à la famille des Stylonuridae (qui sera plus tard relevée pour devenir le sous-ordre Stylonurina, à ne pas confondre avec la famille des Euryptérides Stylonuridae de la classification contemporaine)[9].
Le paléontologue anglais Charles D. Waterston est le premier à suggérer, dans un article de 1958, que C. scouleri ne devrait peut-être pas être considéré comme appartenant au genre Campylocephalus. Il y remarque que, bien que l'anatomie dorsale du prosome de C. scouleri soit assez proche de celle de C. oculatus, l'espèce type, plusieurs fossiles bien conservés ont, depuis leur regroupement dans le même genre, permis des recherches détaillées sur l'anatomie ventrale et les appendices de C. scouleri, tandis que l'anatomie ventrale et les appendices de C. oculatus demeurent inconnus[7]. Un an plus tard, en 1959, le paléontologue américain Erik Norman Kjellesvig-Waering crée le nouveau genre Hibbertopterus pour y affecter C. scouleri (qui devient donc Hibbertopterus scouleri) rattache les deux genres à la famille des Hibbertopteridae[10].
Décrit par le paléontologue russe Alexey G. Ponomarenko en 1985, C. permianus est à l'origine attribuée au genre Hibbertopterus. Le seul spécimen connu de cette espèce est l'holotype PIN N1209/2, constitué d'une carapace incomplète. Mais Ponomarenko parvient, sur cette seule base, à identifier plusieurs caractères qui le distingue des autres espèces d'Hibbertopterus. Parmi ceux-ci figurent en particulier la position postérieure des yeux latéraux et le fait que ces yeux ne soient pas de forme circulaire. En 2012, le paléontologue américain James C. Lamsdell démontre que ces caractères uniques sont en fait des caractéristiques diagnostiques du genre Campylocephalus et reclasse ainsi l'espèce dans son genre contemporain. Bien que Ponomarenko fasse mention de plusieurs caractéristiques qui distinguent également C. permianus des espèces alors attribuées à Campylocephalus, y compris une forme de carapace différente et un épaississement de l'exosquelette autour des yeux, Lamsdell établit que ces distinctions ne sont pas valides. Selon Lamsdell, les spécimens de l'espèce type C. oculatus ne sont pas suffisamment bien conservés pour pouvoir déterminer la structure précise des yeux, et, parce que les fossiles de sa carapace sont soit aplatis soit incomplets, sa forme ne peut pas être déterminée avec une précision totale[1].
Classification
Campylocephalus est classé dans la famille des Hibbertopteridae, une famille d'Euryptérides au appartenant à la super-famille des Mycteropoidea, aux côtés des genres Hibbertopterus et Vernonopterus. Le genre contient trois espèces : C. oculatus et C. permianus du Permien de Russie et C. salmi du Carbonifère de la République tchèque. La classification de C. salmi est considérée comme encore incertaine[10] du fait de l'état fragmentaire de ses fossiles (ce qui est aussi le cas des autres espèces du genre) et de certaines caractéristiques uniques dans le genre (par exemple les protubérances de différentes tailles répartis sur l'exosquelette[5]).
Les Hibbertoptérides forment un groupe de grands Myctéropoïdes caractérisés par leur large prosome, un telson hastaté (c'est à dire en forme de glaive, une épée romaine) présentant des carènes appariées sur la face ventrale, une ornementation composée d'écailles ou d'autres structures similaires sur l'exosquelette, une quatrième paire d'appendices équipée d'épines, les tergites les plus postérieures de l'abdomen présentant des écailles en forme de langue près de leurs bords, et enfin des lobes positionnés postérolatéralement (des deux côtés sur la face postérieure) sur le prosome[11].
Les caractéristiques de Campylocephalus et de Vernonopterus montrent clairement que les deux genres représentent des Euryptérides Hibbertoptérides, mais la nature incomplète de tous les spécimens fossiles qui leur sont attribués rend difficile toute étude plus approfondie des relations phylogénétiques précises au sein des Hibbertopteridae. Les deux genres pourraient même représenter des synonymes d'Hibbertopterus, mais, là encore, la nature très incomplète des fossiles fait que cette hypothèse est impossible à confirmer[11].
Le cladogramme ci-dessous est issu de Lamsdell (2012)[1], réduit pour ne figurer que la super-famille des Mycteropoidea. Il reprend l'hypothèse de Lamsdell d'une affectation du genre Hastimima à la famille des Myctéropides, bien que le consensus scientifique demeure l'affectation à la famille des Hibbertoptérides[10].
| Mycteropoidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||
Les Hibbertoptérides tels que Campylocephalus avaient recours à une méthode d'alimentation par balayage : ils utilisaient leurs appendices prosomaux tournés vers l'avant et équipés d'épines pour ratisser le substrat de leur milieu de vie. Cette stratégie alimentaire a été utilisée par de nombreux genres du sous-ordre des Stylonurina, mais c'est chez les Hibbertoptérides qu'elle apparaît le plus développée : ces derniers possèdent en effet des lames sur les deuxième, troisième et quatrième paires d'appendices. Chez certaines espèces du genre Hibbertopterus, étroitement apparenté à Campylocephalus, ces lames sont spécialisées en rachis, en forme de peigne, capables de piéger des petites proies et d'autres particules alimentaires organiques[11].
L'étude d'empreintes d'Hibbertopterus découvertes en Ecosse semble montrer que les Hibbertoptérides étaient capables de marcher sur la terre ferme, au moins pendant de courtes périodes. On suppose qu'ils étaient lents compte tenu de leur taille et de leur aspect massif. Les traces découvertes semblent indiquer qu'ils se déplaçaient pesamment d'un mouvement saccadé et trainant, le ventre caréné et le telson laissant une rainure centrale sur le sol derrière eux[12]. Certaines études suggèrent que les Euryptérides possédaient un double système respiratoire, ce qui aurait permis ces déambulations terrestres occasionnelles[13].
Paléoécologie
C. salmi a été découvert dans la formation d'Ostrava en République tchèque. Il aurait vécu à l'époque Amsberguienne (il y a 326,4 à 318,1 millions d'années) durant le Carbonifère. Les dépôts où les fossiles de cet Euryptéride ont été trouvés se sont formés au fond d'un ancien lac[14]. Dans la même formation, on a découvert des araignées du genre Eophrynus, de l'ordre éteint des Arachnides Trigonotarbida[14] et de nombreux Anthozoaires (une classe taxonomique qui compte des animaux tels que les coraux et les anémones de mer)[15] contemporains de C. salmi.
La paléoécologie de l'espèce type C. oculatus est difficile à appréhender car le lieu de découverte et la datation du spécimen fossile restent sujet à controverse. La plupart des compte-rendu font état d'une découverte du fossile en un lieu appelé Dourasovo en Russie[10] - [13] et le font remonter à l'époque Guadalupienne (il y a 272,3 à 259,8 millions d'années) au Permien[13].
L'espèce C. permianus est l'une des dernières espèces d'Euryptérides survivantes connues. Le seul fossile représentant l'espèce a été découvert en République socialiste soviétique autonome des Komis à l'époque de l'Union soviétique (la République Komi de Russie moderne) dans des gisements datant approximativement de l'extinction du Permien-Trias il y a 251,9 millions d'années[13]. Au Permien, Komi était probablement une région côtière du supercontinent Pangée. Bien qu'il ne soit fait mention d'aucun autre fossile trouvé dans le même gisement, d'autres formes de vie du Permien ont été découvertes dans la région comme des Bryozoaires[16] et des bivalves[17].
Au début du Permien, les Euryptérides étaient déjà en déclin eu égard à la taille et à la diversité des populations connues antérieurement. Le groupe était l'un des nombreux fortement touchés par l'extinction du Dévonien supérieur qui a vu l'extinction de tous les genres d'Euryptérines (les Euryptérides à palettes natatoires) à l'exception d'Adelophthalmus. Sur les 16 familles d'Euryptérides qui ont vécu au début du Dévonien, seules trois ont survécu et persisté jusqu'au Carbonifère, qui toutes ne comptaient que des espèces non marines[18]. Au Permien, seuls quatre genres étaient encore vivants : Adelophthalmus (un Adelophthalmide), Hastimima et Woodwardopterus (des Myctéroptides), et Campylocephalus lui-même. Adelophthalmus et Hastimima se sont éteints au Permien inférieur, ce qui a fait de Campylocephalus l'un des deux seuls genres vivants d'Euryptérides pendant plus de 20 millions d'années jusqu'à sa propre extinction lors de l'extinction du Permien-Trias[13] - [19] - [20]. Woodwardopterus a également disparu à cette époque[21].
Liens externes
- Ressources relatives au vivant :
Notes et références
- (en) James Lamsdell, « Redescription of Drepanopterus pentlandicus Laurie, 1892, the earliest known mycteropoid (Chelicerata: Eurypterida) from the early Silurian (Llandovery) of the Pentland Hills, Scotland », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 103, , p. 77–103 (DOI 10.1017/S1755691012000072, S2CID 84151264, lire en ligne).
- (en) Leif Størmer, Part P Arthropoda 2, Chelicerata, coll. « Treatise on Invertebrate Paleontology », , 39 p., « Merostomata ».
- (en) James C. Lamsdell et Simon J. Braddy, « Cope's rule and Romer's theory: patterns of diversity and gigantism in eurypterids and Palaeozoic vertebrates », Biology Letters, vol. 6, no 2, , p. 265–269 (ISSN 1744-9561, PMID 19828493, PMCID 2865068, DOI 10.1098/rsbl.2009.0700, lire en ligne).
- Edouard D'Eichwald, Lethaea rossica ou Paléontologie de la Russie, vol. 1, , 1360 p. (lire en ligne), « 2 ».
- (de) Dionýs Štúr, Die Culmflora der Ostrauer und Waldenburger Schichten, vol. 8, Hölder (no 2), , 449–450 p. (lire en ligne).
- (de) Stepan Kutorga, Beitrag zur Kenntniss der organischen Ueberreste des Kupfersandsteins am westlichen Abhange des Urals, Mineralogischen Gesellschaft, (lire en ligne).
- (en) Charles D. Waterston, « XII.—The Scottish Carboniferous Eurypterida* », Earth and Environmental Science Transactions of the Royal Society of Edinburgh, vol. 63, no 2, , p. 265–288 (ISSN 2053-5945, DOI 10.1017/S0080456800009492, S2CID 130625350).
- (en) International Commission on Zoological Nomenclature, Opinions and declarations rendered by the International Commission on Zoological Nomenclature, vol. 1B, Londres, International Trust for Zoological Nomenclature, (lire en ligne).
- (en) Leif Størmer, « A New Eurypterid from the Ordovician of Montgomeryshire, Wales », Geological Magazine, vol. 88, no 6, , p. 409–422 (ISSN 1469-5081, DOI 10.1017/S001675680006996X, Bibcode 1951GeoM...88..409S, S2CID 129426407).
- (en) J. A. Dunlop, D. Penney et D. Jekel, World Spider Catalog : A summary list of fossil spiders and their relatives, Musée d'histoire naturelle de Berne, (lire en ligne).
- (en) James C. Lamsdell, Simon J. Braddy et O. Erik Tetlie, « The systematics and phylogeny of the Stylonurina (Arthropoda: Chelicerata: Eurypterida) », Journal of Systematic Palaeontology, vol. 8, no 1, , p. 49–61 (DOI 10.1080/14772011003603564, S2CID 85398946).
- (en) Robert Roy Britt, « Giant Water Scorpion Walked on Land », Live Science, (lire en ligne).
- (en) O. Erik Tetlie, « Distribution and dispersal history of Eurypterida (Chelicerata) », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 252, nos 3–4, , p. 557–574 (DOI 10.1016/j.palaeo.2007.05.011).
- (en) « Fossilworks - Hermenegilde Shaft, XII Jaklowetzer seam, Ostrava (Carboniferous of Czech Republic) », fossilworks.org (consulté le ).
- (en) « Fossilworks - Sosnica Mine, Gliwice, Upper Roemer Bed (Carboniferous of Poland) », fossilworks.org (consulté le ).
- (en) « Fossilworks - Piles Well, Komi Region (Permian of Russian Federation) », fossilworks.org (consulté le ).
- (en) « Fossilworks - Vym' River, Opposite mouth of Koin River, Komi Region (Permian of Russian Federation) », fossilworks.org (consulté le )
- (en) Anthony Hallam et Paul B. Wignall, Mass Extinctions and Their Aftermath, Oxford University Press, (ISBN 978-0198549161), p. 70.
- (en) David White, « The Flora of the Hermit Shale in the Grand Canyon, Arizona », Proceedings of the National Academy of Sciences of the United States of America, vol. 13, no 8, , p. 574–575 (ISSN 0027-8424, PMID 16587225, PMCID 1085121, DOI 10.1073/pnas.13.8.574, Bibcode 1927PNAS...13..574W).
- (en) Barry S. Kues et Kenneth K. Kietzke, « A Large Assemblage of a New Eurypterid from the Red Tanks Member, Madera Formation (Late Pennsylvanian-Early Permian) of New Mexico », Journal of Paleontology, vol. 55, no 4, , p. 709–729 (ISSN 0022-3360, JSTOR 1304420).
- (en) Markus J. Poschmann et Andrew Rozefelds, « The last eurypterid – a southern high-latitude record of sweep-feeding sea scorpion from Australia constrains the timing of their extinction », Historical Biology, vol. 33, no 12, , p. 121–138 (DOI 10.1080/08912963.2021.1998033, S2CID 252467508, lire en ligne).