Australobarbarus
Australobarbarus est un genre éteint et fossile de thérapsides dicynodontes ayant vécu au début du Permien supérieur (Wuchiapingien inférieur) dans ce qui est aujourd’hui l’est de la Russie européenne. Le genre est représenté par deux espèces, A. kotelnitshi et A. platycephalus qui ont été décrites et nommées en 2000 par Andrey A. Kurkin. Il est étroitement apparenté au genre Tropidostoma trouvé en Afrique du Sud. Comme chez ce dernier, il montre des caractères primitifs comme la rétention d'une postcanine et de quelques dents minuscules sur la mâchoire inférieure.
Espèces de rang inférieur
- A. kotelnitshi Kurkin, 2000
- A. platycephalus Kurkin, 2000
Étymologie
Le nom de genre vient du Latin austral « sud » et barbarus « étranger » et fait référence à l’appartenance de ce taxon à une famille de dicynodontes surtout connus en Afrique australe et orientale, les Oudenodontidae[note 1], dont Australobarbarus est l’un des rares représentants dans l’hémisphère nord[1].
Description
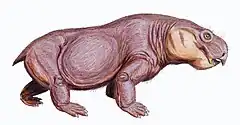
Australobarbarus est un dicynodonte de taille moyenne avec une longueur crânienne de 15 à 20 cm. Le museau présente une crête médiale s’étendant du bord antérieur du prémaxillaire jusqu’au niveau du centre de l’orbite. La mâchoire supérieure possède des excroissances en forme de canines. Ces processus caniniformes sont dirigées ventralement ou antéroventralement selon l’espèce. Des renflements sur la face latérale des maxillaires correspondent à ces processus. Sur le côté postéro-interne de la surface palatine de chaque processus caniniformes, il y a une grande alvéole d’une postcanine située symétriquement. La région pariétale est approximativement de même largeur que la région interorbitaire. Selon l’espèce, les postorbitaires sont contiguës sur la ligne médiane et forment une courte crête, ou sont séparées par un étroit sillon où les pariétaux sont exposés. Les postfrontaux sont petits et coincés entre les frontaux et les postorbitaires. Les palatins sont de taille moyenne, sans contact avec le prémaxillaire, et séparés de ce dernier par des rebords étroits des maxillaires. L’ectoptérygoïde est grand et sépare le maxillaire du ptérygoïde. Les os du palais sont situés dans le même plan que le ptérygoïde. Le bord antérieur des choanes est formé par le prémaxillaire. Les branches antérieure (maxillaire) et postérieure (carré) des ptérygoïdes dessinent une forme en X. Les rameaux antérieurs des ptérygoïdes sont en forme de lyre. L’occiput est large, bas et sensiblement incliné vers l’avant. Les surfaces articulaires des carrés sont situées au niveau du plan de l’occiput. Le basioccipital est dépourvu de crête intertuberculaire. Le prémaxillaire et les ptérygoïdes sont dans le même plan, ce qui suggère une posture naturelle de la tête horizontale. Le foramen magnum est exceptionnellement rétréci, faiblement élargi ventralement et étendu verticalement. Le dentaire est long, et possède une symphyse relativement haute et un plateau dentaire bien développé. Les dents inférieures sont situées près du bord interne de la mâchoire. Les couronnes sont hautes, rondes en coupe transversale, et portent des dentelures sur les bords antérieur et postérieur[1] - [2]
Espèces
Les deux espèces d’Australobarbarus diffèrent essentiellement par la structure des régions préorbitaires et pariétales.
- Australobarbarus kotelnitshi
Le crâne d’Australobarbarus kotelnitschi est subtriangulaire en vue dorsale. Les orbites sont principalement orientées vers l’avant. Les processus caniniformes du maxillaire sont dirigées presque totalement vers le bas, formant un coude distinct par rapport à l’arcade zygomatique. Le tiers postérieur du processus caniniforme est situé sous l’orbite et le bord antérieur s’étend jusqu’à la limite postérieure des narines, alors que chez A. platycephalus, le bord postérieur de ce processus est sous la partie antérieure du bord orbitaire et le bord antérieur se termine au niveau du centre des narines. Le bord postérieur des processus caniniformes est aiguisé. Les narines sont relativement grandes et occupent une position élevée. Dorsalement, elles sont limitées par une excroissance bien développée de l’os nasal qui les surplombe comme une coiffe. Sur une grande partie de la surface latérale des processus caniniformes, on distingue de nombreux foramens relativement larges assurant le passage des vaisseaux sanguins ; cette partie du museau était probablement fortement kératinisé. En général, la structure et la préservation des os indiquent que la kératinisation s'est développée sur la majeure partie du museau, y compris la surface palatine du prémaxillaire et la plupart des palatins. Entre les arcs postorbitaires, il y a une dépression triangulaire bien prononcée, dont la partie supérieure porte un très petit foramen pariétal. Les postorbitaires se rejoignent derrière le foramen pariétal et forment une crête. Le foramen posttemporal est exceptionnellement en forme de fente. La mandibule est relativement basse et porte un plateau dentaire et des rainures sur les branches de la mâchoire. La symphyse est bien développée. La surface articulaire de l’articulaire est à double articulation ; médialement, le condyle interne du carré embrasse une lame originelle de l'articulaire. Les dents inférieures sont relativement petites (sur le spécimen KPM, n° 18/92, la mâchoire inférieure mesure 190 mm de long, les couronnes dentaires mesurent 4–6 mm de haut et 1,5–3 mm de diamètre) et forment une seule rangée au niveau de la bordure médiale de la mâchoire, alors que les surfaces supérieure et latérale de la mandibule étaient probablement kératinisées. Les dents sont situées dans un petit segment allant du tiers postérieur de la symphyse jusqu’au bord antérieur des rainures sur la surface dorsale de la mâchoire. La rangée se compose de trois à cinq dents de différentes tailles. Sur la branche mandibulaire droite du spécimen KPM, no. 18/92, une couronne d'une dent inférieure est presque entièrement conservée. La couronne est haute, étroite, presque cylindrique et s'effile progressivement vers le haut. La dent est ronde en coupe transversale et porte sept denticules distincts dont la taille diminue progressivement ventralement sur les côtés antérieur et postérieur de la couronne. À cet égard, la structure de cette couronne est similaire à celle des postcanines du dinocéphale Estemmenosuchus uralensis où les dents portent des denticules isolés et peu fréquents plutôt que des dentelures caractéristiques de formes prédatrices que sont les thériodontes[1] - [2].
- Australobarbarus platycephalus
Australobarbarus platycephalus se distingue principalement de l’espèce type par l’anatomie de sa région antéorbitaire et temporale. Contrairement aux processus caniniformes d’A. kotelnitshi qui sont dirigées vers le bas, ceux d’A. platycephalus sont dirigées principalement vers l’avant et forment un prolongement de la ligne de l’arcade zygomatique. Ces processus, nettement moins développés que chez A. kotelnitshi, sont situés plus près de la pointe du museau. Chez A. platycephalus, le bord postérieur de ces processus est sous la partie antérieure du bord orbitaire, et le bord antérieur se termine au niveau du centre des narines, alors que chez A. kotelnitshi, le tiers postérieur de ces processus est situé sous l'orbite et le bord antérieur s'étend jusqu'à la limite postérieure des narines. Les orbites sont principalement tournées vers l’avant, mais sont également plus ouvertes vers le haut que chez l’espèce type. En vue dorsale, la distance entre l’arc post-orbitaire et le bord orbitaire antérieur est de 35 mm, alors que chez le spécimen type d’A. kotelnitshi, avec une plus grande longueur de crâne, cette distance est seulement de 25 mm. La partie frontale du crâne est sensiblement plus large. Chez A. kotelnitshi, la distance entre les orbites est de 26 mm, alors que chez A. platycephalus avec une longueur de crâne plus petite, cette distance est de 35 mm. A. platycephalus montre une région pariétale nettement plus large que chez l'espèce type. Les postorbitaires ne se rejoignent pas sur la ligne médiane, exposant ainsi les pariétaux. Un grand foramen pariétal ovale est présent. Une dépression bien prononcée est située en avant du foramen pariétal, comme chez A. kotelnitshi ; cependant, chez ce dernier, il est subtriangulaire et formé presque entièrement par les frontaux (seule la partie postérieure de la dépression est située sur un très petit prépariétal), alors que chez A. platycephalus, la dépression est trapézoïdale en raison d'une région pariétale plus large, et la partie postéro-médiale est formée par le prépariétal. Le prépariétal est sensiblement plus grand et se rétrécit vers l'avant ; le bord antérieur est situé au niveau de la bordure postérieure de l’orbite. La surface palatine est sensiblement plus concave et les deux crêtes palatines antérieures sur la face ventrale du prémaxillaire s'étendent jusqu'à l'extrémité du museau où chacune forme un denticule bien prononcé séparé de son homologue par une incisure superficielle. L'autre distinction substantielle est observée dans la forme du squamosal, à savoir dans la partie temporale de cet os. Chez A. kotelnitshi, l'aile temporale du squamosal forme une surface large et presque plate, alors que chez A. platycephalus, cette aile a une surface concave légèrement effilée vers l'arrière. L'occiput est de forme subtriangulaire. Un spécimen de cette espèce quelque peu écrasé latéralement se distingue par la présence de grandes défenses à section arrondie dans la mâchoire supérieure. C'est le seul spécimen relativement bien conservé d’A. platycephalus préservé avec des défenses. Chez le genre apparenté Tropidostoma on observe deux formes crâniennes, l'une étant une forme robuste avec un museau haut et de grandes défenses et l'autre plus gracile avec un museau bas et des défenses petites ou inexistantes. Ces différences ont été interprété comme un exemple de dimorphisme sexuel. Un tel dimorphisme n’a toutefois pas encore été clairement démontrée chez Australobarbarus[1].
Répartition géographique et stratigraphique
Les fossiles des deux espèces d’Australobarbarus furent découverts sur le site de Port Kotel’nich, sur les berges de la rivière Viatka, dans l’oblast de Kirov en Russie européenne[1]. Ce site, qui appartient à la faune de Kotelnich, est séparé de la section principale de Kotelnich par de mauvaises conditions d’affleurement rendant leur corrélation difficile. Les couches géologiques affleurant à Kotelnich ont été divisées en cinq membres stratigraphiques, du plus ancien au plus récent, les membres de Vanyushonki, Boroviki, Shestakovy, Chizi, et Sokol’ya Gora[3]. Arefiev et des collègues ont reconsidéré ces cinq membres comme quatre couches distinctes (le membre de Chizi étant fusionné avec la couche de Sokol’ya Gora) au sein d’un membre unique, le Membre de Kalininskaya, ce dernier constituant la partie supérieure de la formation de Kotel’nich[4]. La grande majorité des taxons de la faune de Kotelnich proviennent du membre ou de la couche de Vanyushonki[3] - [4]. Selon Areviev et des collègues, le site de Port Kotel’nich se trouverait plus haut dans la succession et appartiendrait à la couche de Shestakovy[4]. En raison de la coexistence de nombreux thérapsides basaux (les gorgonopsiens Nochnitsa et Viatkogorgon, les thérocéphales Perplexisaurus et Gorynychus, l’anomodonte Suminia) avec des thérocéphales plus dérivées (le Whaitsiidae Viatkosuchus et le Baurioidea Karenites), la faune du membre de Vanyushonki a’été mise en corrélation avec la sous-zone à Lycosuchus – Eunotosaurus (= ancienne zone à Pristerognathus) de la Zone d’association à Endothiodon du bassin du Karoo en Afrique du Sud[5]. La base de la sous-zone à Lycosuchus – Eunotosaurus est relativement bien contrainte à environ 260 millions d’années par un âge radiométrique de 260,259 ± 0081 Ma à partir de la base du membre de Poortjie de la formation de Teekloof[6] - [5]. La limite supérieure de la sous-zone n’est pas connue avec certitude mais se situerai entre 259 et 258 Ma, lui conférant un âge englobant le Capitanien terminal et le Wuchiapingien basal. Sur la base de ces corrélations, la faune de Kotelnich pourrait avoir un âge Capitanien terminal et/ou Wuchiapingien basal. La faune de la couche plus récente de Shestakovy, qui contient Australobarbarus, date probablement du Wuchiapingien inférieur[5].
Liens externes
- Ressources relatives au vivant :
Notes et références
Notes
- Australobarbarus fut attribué aux Cryptodontidae par Kurkin en 2000, mais ce nom de famille fut remplacé par Oudenodontidae (Kammerer & Angielczyk 2009 ; Kurkin 2010)
Références
- (en) A.A. Kurkin, « New Dicynodonts from the Upper Permian of the Vyatka Basin », Paleontological Journal, vol. 34, no 2, , p. 203-210
- (en) A.A. Kurkin, « Late Permian Dicynodonts of Eastern Europe », Paleontological Journal, vol. 44, no 6, , p. 672-681 (DOI 10.1134/S0031030110060092)
- (en) M.J. Benton, A.J. Newell, A.Yu. Khlyupin, I.S. Shumov, Price et A.A. Kurkin, « Preservation of exceptional vertebrate assemblages in Middle Permian fluviolacustrine mudstones of Kotel’nich, Russia: stratigraphy, sedimentology, and taphonomy », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 319-320, , p. 58-83 (DOI 10.1016/j.palaeo.2012.01.005)
- (en) M.P. Areviev, V.K. Golubev, Yu.P. Balabanov, E.V. Karasev, A.V. Minikh, M.G. Minikh, I.I. Molostovskaya, O.P. Yaroshenko et M.A. Zhokina-Naumcheva, « Part I. Middle Permian – Lower Triassic continental sections of the Sukhona, Malaya Severnaya Dvina, Kichmenga and Vyatka rivers », dans Arefiev, M.P., Golubev, V.K., Balabanov, Yu.P., Karasev, E.V., Minikh, A.V., Minikh, M.G., Molostovskaya, I.I., Yaroshenko, O.P.& Zhokina-Naumcheva, M.A., Type and reference sections of the Permian-Triassic continental sequences of the East European Platform: main isotope, magnetic, and biotic events, Moscow, Borissiak Paleontological Institute of RAS, (ISBN 978-5-903825-34-9), p. 50–53
- (en) M.O. Day et R.M.H. Smith, « Biostratigraphy of the Endothiodon Assemblage Zone (Beaufort Group, Karoo Supergroup), South Africa », South African Journal of Geology, vol. 123, no 2, , p. 165-180 (DOI 10.25131/sajg.123.0011)
- (en) M.O. Day, J. Ramenazi, S.A. Bowring, P.M. Sadler, D.H. Erwin, F. Abdala et B.S. Rubidge, « When and how did the terrestrial mid-Permian mass extinction occur? Evidence from the tetrapod record of the Karoo Basin, South Africa », Proceedings of the Royal Society B, vol. 282, no 1811, (DOI 10.1098/rspb.2015.0834)