Anatomie de type kranz
En botanique, l'anatomie de type kranz, ou structure kranz (du terme allemand « Kranz » signifiant « couronne »), est une structure anatomique particulière des feuilles qui se rencontre chez la plupart des plantes en C4. Elle se caractérise par la présence de deux anneaux de cellules concentriques entourant les faisceaux vasculaires : l'anneau interne, composé de cellules serrées contenant des chloroplastes de grande taille, riches en amidon dépourvus de grana, forme la gaine fasciculaire, l'anneau externe composé de cellules plus lâches du mésophylle[1] - [2]. Les cellules du mésophylle présentent également des chloroplastes, mais ceux-ci ne synthétisent pas d'amidon car ils n'effectuent pas le cycle de Calvin réservé aux chloroplastes de la gaine[3]. Par conséquent, les chloroplastes sont dits « dimorphes ».
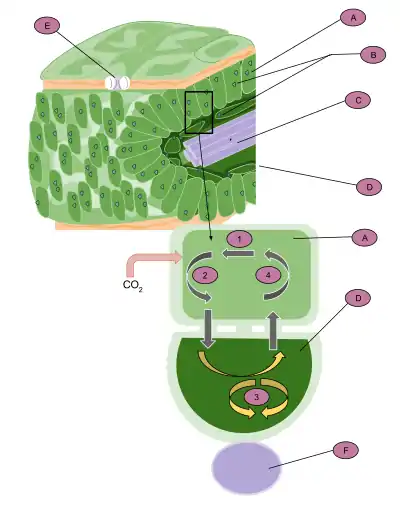
A : cellule du mésophylle, B : chloroplaste, C : tissu vasculaire, D : cellule de la gaine fasciculaire, E : stomate, F : tissu vasculaire : fournit une source continue d'eau. 1) Le carbone est fixé pour produire de l'oxaloacétate par la carboxylase PEP.
2) La molécule à quatre carbones quitte ensuite la cellule et entre dans les chloroplastes des cellules de la gaine fasciculaire.
3) Elle est ensuite décomposée libérant du dioxyde de carbone et produisant du pyruvate. Le dioxyde de carbone se combine avec le bisphosphate de ribulose et passe au cycle de Calvin.
4) le pyruvate rentre dans la cellule du mésophylle et réagit ensuite avec l'ATP pour produire le composé de départ du cycle en C4.
La fonction principale de l'anatomie kranz est de fournir un site dans lequel le CO2 peut être concentré autour de la rubisco (ribulose-1,5-bisphosphate carboxylase/oxygénase), évitant ainsi la photorespiration. Afin de maintenir une concentration de CO2 dans la gaine fasciculaire significativement plus élevée que dans le mésophylle, la couche limite du kranz a une faible conductance au CO2, propriété qui peut être améliorée par la présence de subérine[4]. En augmentant le cycle interne du dioxyde de carbone, cette anatomie pourrait entraîner une réduction de la sensibilité des plantes à la photo-inhibition[5]. En effet, la couche de cellules du mésophylle, située dans la partie la plus externe, capture les molécules de CO2 et les concentre en les convertissant en molécules de sucre à quatre atomes de carbone, pour les céder aux cellules qui composent la couche interne[6].
Cette voie de formation des sucres du mésophylle est appelée « voie Hatch Slack ». Au cours de la première étape, il se forme de l'acide oxaloacétique par réaction du dioxyde de carbone avec le phosphoénolpyruvate. À partir de là, se produit une transformation dont le résultat diffère selon le type de plante. Le produit passe dans les cellules de la gaine par l'intermédiaire des plasmodesmes et par une série de transformations, du dioxyde de carbone est libéré et entre dans le cycle de Calvin[7].

Bien que la plupart des plantes en C4 présentent une anatomie de type kranz, il existe cependant quelques espèces qui opèrent un cycle C4 limité sans aucun tissu distinct de gaine fasciculaire.
Des espèces de la sous-famille des Chenopodioideae (Suaeda aralocaspica, Bienertia cycloptera, Bienertia sinuspersici et Bienertia kavirense), plantes terrestres poussant dans des dépressions sèches et salées des déserts du Moyen-Orient, fonctionnent avec des mécanismes de concentration du CO2 à cellule unique en C4. Ces mécanismes sont uniques parmi les mécanismes en C4 connus[8] - [9] - [10] - [11]
Bien que la cytologie des deux genres (Suaeda et Bienertia) soient différente, le principe de base est que les vacuoles remplies de liquide sont utilisées pour diviser la cellule en deux zones distinctes. Les enzymes de carboxylation dans le cytosol peuvent donc être séparées dans les chloroplastes des enzymes décarboxylases et rubisco, et une barrière diffusive peut être établie entre les chloroplastes (qui contiennent de la rubisco) et le cytosol. Cela permet d'établir une zone de type gaine fasciculaire et une zone de type mésophylle au sein d'une même cellule. Bien que cela permette un cycle C4 limité, celui-ci est relativement inefficace, avec de nombreuses fuites de CO2 autour de la rubisco.
On a également montré la possibilité de photosynthèse en C4 inductible chez des macrophytes, hydrilla verticillata, dans des conditions chaudes, bien que le mécanisme qui minimise la fuite de CO2 à partir de la rubisco est actuellement incertain[12].
Notes et références
- (es) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en espagnol intitulé « Anatomía Kranz » (voir la liste des auteurs).
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « C4 carbon fixation » (voir la liste des auteurs).
- Daniel Richard, Patrick Chevalet, Sylvie Fournel, Nathalie Giraud, Frédéric Gros, Patrick Laurenti, Fabienne Pradere, Thierry Soubaya, Biologie -Tout le cours en fiches : 300 fiches de cours, 270 QCM et bonus web : Tout le cours en fiches, Dunod, , 768 p. (ISBN 978-2-10-072328-7), p. 220.
- (es) Peter H. Raven, Ray F. Evert et Susan E. Eichhorn, « 22 - El vástago: Estructura primaria y desarrollo », dans Biología de las plantas, vol. 2, Reverte, , 432 p. (ISBN 9788429118421, lire en ligne), p. 402.
- (es) Rosa Redes Garcia et Margarita Collazo Ortega, Manual de Practicas de Fotosintesis, UNAM, , 160 p. (ISBN 978-970-32-3313-7, lire en ligne), p. 61
- Laetsch (1971) Photosynthesis and Photorespiration, ed. Hatch, Osmond & Slatyer.
- (es) Z. Baruch et M.J. Fischer, « Factores climáticos y de competencia que afectan el desarrollo de la planta en el establecimiento de una pastura », dans Establecimiento y renovación de pasturas: conceptos, experiencias y enfoque de la investigación, Red Internacional de Evaluación de Pastos Tropicales, Sexta Reunión del Comité Asesor, CIAT, , 425 p. (ISBN 9789589183267, lire en ligne), p. 121.
- (es) Centro Internacional de Agricultura Tropical, CIAT report, Université du Texas, Centro Internacional de Agricultura Tropical, (lire en ligne), p. 39
- (es) Francisco José García Breijo, Josefa Roselló Caselles et Pilar Santamarina Siurana, Introducción al funcionamiento de las plantas, Ed. Univ. Politéc. Valencia, , 182 p. (ISBN 978-84-9705-944-2, lire en ligne), p. 32.
- (en) H Freitag et W Stichler, « A remarkable new leaf type with unusual photosynthetic tissue in a central Asiatic genus of Chenopodiaceae », Plant Biol, vol. 2, , p. 154–160 (DOI 10.1055/s-2000-9462).
- (en) Elena Voznesenskaya, Vincent R. Franceschi, Olavi Kiirats, Elena G. Artyusheva, Helmut Freitag et Gerald E. Edwards, « Proof of C4 photosynthesis without Kranz anatomy in Bienertia cycloptera (Chenopodiaceae) », The Plant Journal, vol. 31, no 5, , p. 649–662.
- (en) Hossein Akhani, João Barroca, Nuria Koteeva, Elena Voznesenskaya, Vincent Franceschi, Gerald Edwards et Seyed Mahmood Ghaffari, « Bienertia sinuspersici (Chenopodiaceae): A New Species from Southwest Asia and Discovery of a Third Terrestrial C4 Plant Without Kranz Anatomy », Systematic Botany, vol. 30, no 2, , p. 290–301 (DOI 10.1600/0363644054223684).
- (en) H Akhani, T Chatrenoor, M Dehghani, R Khoshravesh, P. Mahdavi et Z. Matinzadeh, « A new species of Bienertia (Chenopodiaceae) from Iranian salt deserts: a third species of the genus and discovery of a fourth terrestrial C4 plant without Kranz anatomy », Plant Biosystems, vol. 146, , p. 550–559 (DOI 10.1080/11263504.2012.662921).
- (en) A. S. Holaday et G. Bowes, « C4 Acid Metabolism and Dark CO2 Fixation in a Submersed Aquatic Macrophyte (Hydrilla verticillata) », Plant Physiology, vol. 65, no 2, , p. 331–5 (DOI 10.1104/pp.65.2.331, résumé).