Banksia attenuata
Le Banksia côtier (Banksia attenuata), ou Banksia chandelier, est une espèce de plantes arbustives de la famille des Proteaceae. Il peut se présenter sous la forme d'un arbre, pouvant atteindre 10 m de haut, ou d'un arbuste buissonnant dans des zones plus sèches, et il ne dépasse alors pas 0,4 à 2 m de haut. Il a de longues feuilles étroites dentelées et des inflorescences jaune vif, qui sont bien visibles au-dessus du feuillage au printemps et en été. Les fleurs tournent au gris et se fanent lorsque se développent les follicules lignifiés. On le trouve à travers le sud-ouest de l'Australie-Occidentale, du nord du parc national de Kalbarri au Cap Leeuwin au sud, ainsi que dans le parc national de Fitzgerald River.

John Lindley baptise des échantillons rapportés par James Drummond en 1840 Banksia cylindrostachya, mais ils se montrent finalement identiques à l'espèce nommée Banksia attenuata par Robert Brown 30 ans plus tôt en 1810, et cette dernière appellation est donc conservée. Au sein du genre Banksia, la position exacte de B. attenuata n'est pas clairement établie.
Banksia attenuata est pollinisé et constitue une source de nourriture pour une large variété de vertébrés et d'invertébrés pendant l'été. Plusieurs espèces se nourrissant de nectar visitent ses fleurs, comme la souris à miel, qui joue un rôle important dans sa pollinisation. Banksia attenuata a la capacité de repartir depuis son lignotuber après un incendie, ou à partir de bourgeons épicormiques situés le long de son tronc. Cette plante a une espérance de vie pouvant aller jusqu'à 300 ans. Il a été largement planté dans les rues en Australie-Occidentale. Toutefois, du fait de sa grande taille, il ne peut être installé dans de petits jardins. Une forme naine est commercialisée pour pallier cet inconvénient.
Description

Banksia attenuata se présente généralement sous la forme d'un arbre de 10 m de haut. Dans le nord de son aire de répartition, où le climat est nettement plus chaud et sec, il a la forme d'un arbuste de 0,4 à 2 m de haut. On peut voir ces deux formes à proximité d'Hill River, mais autrement elles cohabitent rarement et sont bien distinctes[1]. Dans la Wheatbelt et dans l'est de la chaîne de Stirling, c'est un arbre chétif. Les formes arborées ont un tronc solide, généralement ondulé ou courbé, recouvert d'une écorce friable de 1 à 2 cm d'épaisseur et de couleur orange-grise dehors et marron-rouge à l'intérieur[2]. Il possède la capacité de se régénérer après un incendie grâce à son lignotuber ou ses bourgeons épicormiques que l'on trouve dans son tronc très résistant aux flammes. Il a de longues feuilles étroites de 4 à 27 cm de long pour 0,5 à 1,6 cm de large[3] - [4]. Les bords de la feuille sont marquées de dentelures en forme de V ou de U sur toute la longueur du limbe. Les jeunes feuilles sont d'un gris-vert pâle quand elles émergent à la fin du printemps et en été[5], souvent après la floraison. Les inflorescences jaune vif apparaissent au printemps et en été et mesurent 5 cm de large pour 25 cm de hauteur[2]. Elles sont constituées de plusieurs petites fleurs, une étude menée au mont Adams, à 330 km au nord de Perth, ayant révélé que chaque inflorescence en comptait 1933 (avec un écart-type de 88)[6], tandis qu'une autre conduite dans le parc national de Fitzgerald River a conclu à 1720 (± 76) fleurs par inflorescence.
L'anthèse est séquentielle, c'est-à-dire que toutes les fleurs de l'inflorescence ne fleurissent pas au même moment. Elle dure entre 10 et 20 jours[7]. Souvent vert vif lorsqu'elles n'ont que des bourgeons floraux[8], les inflorescences se trouvent en position terminale, à l'extrémité des branches de trois ans. Elles sont bien visibles au-dessus du feuillage[2]. Les fleurs, une fois ouvertes, dégagent une odeur comparable à un vin du cépage syrah bien poivré[8]. Au bout d'un moment, les inflorescences passent au marron, puis au gris[3] et leurs fleurs se flétrissent. C'est alors que commence à se développer un follicule sombre et poilu, de forme ovale, mesurant 2 à 3,5 cm de long pour 1 à 1,5 cm de haut et 1,4 à 2 cm de large[2]. Toutefois, seul un faible pourcentage (0,1 %) de fleurs se développent pour former des follicules ; le pourcentage lors de l'étude au mont Adams était de 3,6 par inflorescence, avec un écart-type de 1,2[6]. Les follicules mettent entre sept et huit mois pour se développer, entre février et décembre, tandis que la formation des graines se fait sur quatre mois entre septembre et décembre[9].
Taxonomie

Banksia attenuata est collecté et décrit pour la première fois dans la King George Sound en décembre 1801 par Robert Brown, qui publie officiellement sa découverte en 1810. Le nom de l'espèce est le terme latin attenuatus, signifiant « étroit » et faisant référence aux feuilles étroites à leur base[3]. Cette espèce a une taxonomie assez simple, avec seulement deux synonymes et aucune sous-espèce ni variété. Après avoir longuement étudié les variations au sein de l'espèce et observé que les formes arborées et arbustives différaient uniquement par leur taille sans que cela ne justifie de les classer en deux taxa distincts[2], John Lindley fait apparaître en 1840 une éventuelle nouvelle espèce, Banksia cylindrostachya, dans son A Sketch of the Vegetation of the Swan River Colony, mais elle se révèle finalement être un synonyme de B. attenuata[10]. En 1891, la tentative infructueuse d'Otto Kuntze de changer le nom d'espèce Banksia en Sirmuellera laisse à celui-ci le temps de renommer l'espèce en Sirmuellera attenuata, aujourd'hui considéré comme un synonyme de B. attenuata[11]. Les noms vernaculaires qui lui sont attribués incluent « Banksia élancé », « Banksia chandelier », « Banksia côtier » et « Banksia à feuilles rétrécies ». Dans la région de Melville, non loin de Perth, les aborigènes d'Australies l'appellent Piara (ou biara).
La position exacte de Banksia attenuata au sein du genre Banksia n'est pas claire. Quand Carl Meissner publie sa classification des Banksias en 1856, il place B. attenuata dans la section Eubanksia du fait de son inflorescence qui a une forme pointue, et non en dôme, et dans la série Salicinae[12], une série assez importante qui apparait aujourd'hui très hétérogène[2]. Elle est d'ailleurs remise en cause par la classification de George Bentham, qui place lui B. attenuata dans la section Cyrtostylis, regroupant des espèces ne pouvant être facilement incluses dans les autres sections[13].
En 1981, Alexander Segger George publie une classification révisée qui place B. attenuata dans le sous-genre Banksia, s'appuyant encore une fois sur la forme de son inflorescence, dans la section Banksia du fait de son pistil droit et non en crochet, et dans la série Cyrtostylis, série assez hétérogène comportant 12 espèces. Il admet toutefois que ses grands cotylédons émarginés sont assez différents de ceux des autres membres de la série et qu'il a des similarités avec B. elegans, un autre membre de la série qui sort lui aussi un peu du lot. Il remarque également que B. attenuata présente des similitudes avec B. lindleyana et B. media[2].


La classification de George reste d'actualité jusqu'en 1996, quand Kevin Thiele et Pauline Ladiges publient leur propre classification s'appuyant sur une analyse cladistique des caractéristiques morphologiques des différentes espèces. Ils placent B. attenuata dans un large clade comprenant notamment B.ashbyi, mais admettent qu'un travail supplémentaire serait nécessaire pour bien déterminer les relations au sein de ce clade, le laissant comme une incertae sedis (ce qui signifie que sa place exacte n'est pas claire)[14]. Interrogé sur l'analyse qu'il porte sur la classification de Thiele et Ladiges, George a publié une version légèrement retouchée de sa classification de 1981 dans un article de 1999 paru dans la série d'ouvrages Flora of Australia. Cette dernière classification reste la plus approfondie à ce jour. D'après celle-ci, on peut placer B. attenuata de la manière suivante[4] :
Depuis 1998, le botaniste américain Austin Mast a publié les résultats d'une analyse cladistique s'appuyant sur des données issues de séquences d'ADN pour les Banksiinae, qui comprennent le genre Banksia et les Dryandra. Ils proposent une phylogénie bien différente de la classification de George. Banksia attenuata apparait comme une base, très proche d'un clade voisine comprenant notamment B. elegans et du sous-genre monophylétique B. subg. Isostylis[15] - [16] - [17]. Un cône fossile datant de l'Éocène nommé Banksia archaeocarpa et âgé d'environ 50 millions d'années, ressemble à B. attenuata[18].
En 2007, Mast et Thiele réorganise le genre Banksia en le fusionnant avec le genre Dryandra et crée le taxon B. subg. Spathulatae pour les espèces ayant des cotylédons en forme de cuillère ; ainsi B. subg. Banksia est redéfini comme un taxon comportant les espèces n'ayant pas de cotylédons en forme de cuillère. Tout cela laisse présager la publication d'un arrangement complet de la classification de ces espèces une fois que l'étude de l'ADN des espèces du genre Dryandra sera terminée. Même si les changements de nomenclature réalisés par Mast et Thiele sont considérés comme provisoires, on peut observer qu'ils ont classé B. attenuata dans le sous-genre B. subg. Banksia[19].
Répartition géographique et habitat
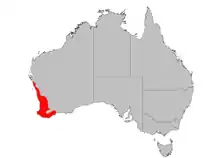
Banksia attenuata est une des espèces de Banksia de l'ouest du pays la plus largement distribuée. Elle est présente dans une large partie du sud-ouest de l'Australie-Occidentale, depuis le parc national de Kalbarri et de la rivière Murchison jusqu'à l'extrémité sud-ouest de l'état à Augusta et au Cap Leeuwin, et à l'est jusqu'au sud du parc national de Fitzgerald River. Son aire de répartition s'étend à l'est jusqu'à Lake Grace, au lac Magenta situé au nord de Jerramungup et à Wongan Hills. On le trouve uniquement dans des sols sableux, sur des sables blancs jaunes ou marron, et sur des sables reposant sur de la latérite ou du calcaire. Il ne pousse pas sur les sols lourds de type argileux. C'est un composant important des forêts d'eucalyptus, en position dominante ou plus secondaire sous ses formes arborées ou arbustives. Au nord de son aire de répartition, il est omniprésent dans les broussailles[5]. Dans les forêts assez ouvertes, on le trouve aux côtés de B. menziesii, B. ilicifolia, B. prionotes, Allocasuarina fraseriana, Eucalyptus marginata, ou E. gomphocephala[2]. Les précipitations annuelles au sein de son habitat varient entre 300 et 900 mm[8].
Écologie
Adaptation aux feux de broussailles

Comme plusieurs autres plantes du sud-ouest de l'Australie, B. attenuata est adapté à un environnement où les feux de broussailles sont courants. Parmi les espèces de Banksia, on observe deux stratégies différentes pour survivre au feu : certaines espèces sont tuées par le feu, mais ce dernier provoque aussi la libération de graines qui permettront la naissance d'une nouvelle génération de plantes, tandis que d'autres espèces survivent aux flammes et repartent à partir de leur lignotuber, ou plus rarement de leurs bourgeons épicormiques, qui sont protégés par une épaisse écorce[20]. Avec ses bourgeons épicormiques et son lignotuber, B. attenuata fait partie de la seconde catégorie.
Cette espèce est faiblement sérotinique, conservant seulement un tiers de ses graines dans une coque de bois, de la même manière que B. hookeriana avec lequel il cohabite sur les dunes de sable de l'Eneabba au nord de Perth. Toutefois, certains de ces follicules ne relâchent pas de graines après un incendie, mais après plusieurs automnes pluvieux successifs[21]. Une expérience simulant un temps humide à la suite d'un incendie a tenté de vérifier cela. Les follicules étaient soumis à deux immersions hebdomadaires dans l'eau après avoir été chauffés à la flamme d'un bec Bunsen à environ 500 à 600 °C pendant deux minutes. On a alors pu observer que plus les follicules étaient soumis longtemps aux immersions, plus il y en avait qui s'ouvraient : environ 40 % des follicules ont relâché leur graine après trois semaines d'immersions répétées, et ce pourcentage augmenta pour atteindre 90 % au bout de 10 semaines. Cela alors que dans un échantillon de follicules conservés au sec moins de 10 % ont libéré leur graine. Cette faculté de l'espèce lui permet de libérer ses graines pendant l'hiver, une période plus humide et donc plus favorable à leur germination qu'un été sec[22].
Dispersion des graines
Après l'ouverture du follicule, la graine et sa membrane ligneuse sont alors soumis aux éléments. Les ailes de la membranes sont hygroscopiques, et elles se mettent en mouvement sous l'action de l'humidité, et se rétractent par temps sec. La succession de périodes humides finit par conduire à l'extraction progressive de la graine[22]. Une fois relâchées, les graines germent à une température située entre 15 et 20 °C, pour optimiser leur chance de se développer avec les pluies d'automne et d'hiver. Toutefois, de nombreuses plantules disparaissent durant les mois d'été chauds et secs[23]. La survie des jeunes plantes de cette espèce est peu élevée par rapport à diverses autres espèces du genre Banksia. Toutefois, la longévité des plantes adultes permet de maintenir la population jusqu'à ce qu'une année favorable permette le développement de plus de nouveaux plants. Une fois arrivées à maturité, les plantes ne risquent pratiquement plus de périr, et peuvent vivre jusqu'à 300 ans ou plus[24]. En analysant le lit de semence et l'historique des résultats d'études menées sur les 15 dernières années dans la région d'Eneabba, on s'est aperçu que B. attenuata devient plus abondant avec un intervalle entre les feux de forêt de 6 à 20 ans, et notamment si ces intervalles sont de 10 à 12 ans, alors que B. hookeriana et B. prionotes, qui s'appuient sur une forte production de graines plutôt que sur la possibilité de repousse de leur lignotuber en cas d'incendie, deviennent abondants dans les secteurs où les feux ont été plus espacés dans le temps. Face à ces principaux concurrents, B. attenuata va prendre l'ascendant dans des zones où les feux sont espacés de 8 à 10 ou 11 ans, mais pour des intervalles plus importants il va être dépassé par d'autres espèces telles que B. hookeriana. La grande variabilité qu'on observe sur le terrain quant à cet intervalle entre deux feux de forêt permet aux trois espèces citées de cohabiter. Des conditions météorologiques extrêmes, bonnes ou mauvaises, favorisent B. attenuata par rapport aux deux autres espèces[25].
Les graines de Banksia attenuata sont relativement lourdes, mais disposent tout de même d'une forte capacité de dispersion sur de longues distances. Une étude génétique des populations menée à Eneabba a montré que plus de 5 % des plantes provenaient de graines émises à plus de 2,6 km (des résultats similaires à Banksia hookeriana, dont la graine pèse moitié moins lourd). Le mécanisme de dispersion n'est pas très bien connu, bien que Byron Lamont pense que le cacatoès à rectrices blanches (Calyptorhynchus latirostris) est un important vecteur de dispersion pour ces graines ; cette espèce recherche les cônes de Banksia attenuata après les feux de broussailles, peut-être du fait de la taille importante des graines et de la possibilité de trouver des insectes réfugiés dans les cônes, ce qui les rend plus nutritifs[26]. La plante refleurit un ou deux ans après l'incendie[5].
Reproduction

Comme plusieurs autres membres de la famille des Proteaceae, Banksia attenuata est une espèce allogame. Ainsi, les inflorescences nécessitent l'intervention de pollinisateurs extérieurs pour produire des graines. Une étude génétique de graines collectées près de Jandakot publiée en 1980 a clairement établi cette allogamie[27]. Une étude de terrain a été menée dans le parc national de Fitzgerald River, au cours de laquelle des inflorescences ont été enfermées dans de petites cages aux grilles suffisamment fines pour empêcher les abeilles de passer a montré que même de tout petits animaux étaient capables d'assurer la pollinisation de l'espèce[7]. Les fleurs de Banksia attenuata sont notamment butinées par l'abeille plâtrière Hylaeus globuliferus et des abeilles appartenant au genre Euhesma[28]. D'autres pollinisateurs ont pu être ponctuellement observés comme des fourmis ou des libellules[5]. Une étude portant sur la population d'invertébrés évoluant dans la canopée des forêts de Banksia a montré une prédominance des acariens, des coléoptères et des guêpes (hyménoptères). Ces trois ordres sont les plus communs sur le B. attenuata, avec les thrips (thysanoptères). Globalement, on trouve tout de même moins d'invertébrés sur les espèces de Banksia que sur d'autres plantes du fait de la présence d'oiseaux insectivores[29].
Plusieurs espèces d'oiseaux ont été relevés sur B. attenuata par la National Banksia Atlas Survey, comme le Méliphage de Nouvelle-Hollande (Phylidonyris novaehollandiae), le Méliphage brunâtre (Lichmera indistincta), le Méliphage chanteur (Lichenostomus virescens), le Méliphage festonné (Acanthorhynchus superciliosus), la Perruche à collier jaune (Barnardius zonarius semitorquatus) et le Cacatoès banksien (Calyptorhynchus banksii)[5]. Des cacatoès de couleur noire ont été observés se nourrissant de graines de B. attenuata, mais l'espèce exacte à laquelle ils appartenaient n'a pas pu être identifiée, s'il s'agissait du Cacatoès à rectrices blanches ou du Cacatoès de Baudin (Calyptorhynchus baudinii)[30]. Près de Jandakot, des Cacatoès à rectrices blanches ont été observés triant des inflorescences immatures qui présentaient des signes d'infestation par le Curculionoidea Alphitopis nivea, dont la larve creuse dans les fleurs de Banksia et mange la graine. Les cacatoès extraient la larve et laissent tomber l'inflorescence[31].
Une étude de 1978 menée dans le comté d'Albany a prouvé que la souris à miel (Tarsipes rostratus) jouait un rôle important dans la pollinisation de Banksia attenuata, puisqu'elle se nourrit directement de pollen et boit le nectar. La structure de la fleur est bien adaptée pour recouvrir l'animal de pollen lorsqu'il se nourrit, ce qui n'est pas le cas avec les méliphages dont le bec est trop long pour qu'ils puissent bien jouer un rôle de pollinisateur. Si l'on combine cela avec l'odeur très musquée de la fleur, on peut en conclure que Banksia attenuata est très bien adaptée à la pollinisation par les mammifères[32]. Par ailleurs, Petroc Sumner et son équipe ont étudié les cellules photoréceptrices en cônes de la souris à miel et les ont comparé avec les changements de couleur de B. attenuata. Ils ont découvert que cet animal avait une vision trichromatique (comme les humains et certainement divers marsupiaux), et les scientifiques pensent que ses cônes L l'aident à discerner les fleurs de B. attenuata, tandis que ses cônes M lui permettent de différencier les inflorescences pleines de nectar de celles qui ont déjà été consommées (ce qui est très difficile à faire pour l'homme)[33]. D'autres mammifères participent occasionnellement à la pollinisation de l'espèce. On a ainsi trouvé du pollen très ressemblant à celui de B. attenuata sur des peaux de souris marsupiales et d'opossums pygmées appartenant au genre Cercartetus. Par ailleurs, le Phalanger volant (Petaurus breviceps) a été observé en captivité visitant des inflorescences[32].
17 espèces appartenant à divers ordres de myxomycètes ont été observées sur l'écorce de Banksia attenuata. Neuf d'entre elles appartenaient à l'ordre des Stemonitidae, et les Echinosteliales et Liceida comprenaient une majeure partie des autres. La forte présence des deux premiers ordres cités est peut-être liée à l'acidité de l'écorce. Un autre ordre, celui des Physariida, est inhabituellement très rare, alors que diverses études ont montré son abondance sur l'écorce de diverses espèces d'arbres à travers le monde[34].
Système racinaire

Tous les Banksia ont développé des racines protéoïdes pour s'adapter sols australiens peu fournis en nutriments, et notamment en phosphore[35]. On a trouvé ces racines jusqu'à 15 cm de profondeur dans la région d'Eneabba[36]. La plante développe une quantité de fines racines latérales qui forment une structure semblable à un tapis juste en dessous du sol, et la rendent capable d'extraire des nutriments du sol le plus efficacement possible. Une étude concernant trois espèces cohabitant dans les bois de Banksia du sud-ouest de l'Australie — Banksia menziesii, B. attenuata et B. ilicifolia — a montré que toutes trois développent des racines nouvelles en septembre après les pluies hivernales, et que la population de bactéries associée au système racinaire de B. menziesii diffère de celle associée aux deux autres, et qu'elle change également en fonction de l'âge des racines[37]. Une autre étude portant sur l'architecture racinaire de Banksia hookeriana, B. menziesii et B. attenuata a montré que la structure globale était similaire, avec des « tapis » protéoïdes plus actifs et croissant durant les mois les plus humides (en hiver et au printemps). Les plantes développent également quelques racines profondes qui vont chercher l'eau en profondeur, au niveau de la nappe phréatique, tandis que la racine pivot d'origine a pu disparaître[38]. De la même manière que B. amenziesii, B. attenuata est phréatophyte facultatif. Ces deux espèces sont moins fortement liées à la nappe phréatique que d'autres et peuvent s'établir dans des endroits plus divers comme dans les bois de Banksia environnants, contrairement à B. ilicifolia et B. littoralis[39].
Une étude menée à l'emplacement d'une ancienne mine de sable recolonisée par la végétation au nord de Perth a montré que les espèces à feuilles larges comme B. attenuata et B. hookeriana ont plus de mal à s'installer que les espèces à feuilles étroites comme B. leptophylla. Ceci s'explique certainement par l'importante impédance du sol perturbé[40]. De même, des analyses réalisées sur de vieux plants dans les restes d'un boisement de Banksia de la banlieue de Perth, où se sont fortement développées deux espèces herbacées invasives (Ehrharta calycina et Pelargonium capitatum), ont montré que les feuilles d'origine contenaient des concentrations élevées de phosphore. Bien que les feuilles de B. attenuata ne montrent pas de niveaux de phosphore croissant, elles ont réduit leur teneur en manganèse, un élément absorbé par les racines protéoïdes, dont la formation peut être inhibée par de forts niveaux de phosphore[41].
Dans une étude de 1985 sur des pieds inoculés, Banksia attenuata s'est révélé modérément à très sensible à Phytophthora cinnamomi qui cause la pourriture de ces racines[42], ce qui se confirme lors d'observations sur le terrain[43]. P. cinnamomi se répand de plante en plante via les racines latérales des arbres. Elle avance à la vitesse d'un mètre par an. Les symptômes présentés par les plants de Banksia attenuata touchés sont le jaunissement des feuilles situées à proximité du tronc et la présence de lésions à la base de celui-ci. Les racines habituellement rouges virent au marron[44]. Une étude menée dans une forêt de Banksia attenuata à 400 km au sud-est de Perth sur 16 ans, à la suite d'une vague d'infestation par P. cinnamomi, a montré que B. attenuata parvenait à se maintenir mais que la population avait été grandement diminuée dans les zones les plus touchées[45]. En injectant une solution à base de phosphite dans des troncs de plants de B. attenuata malades, sur le front d'infection d'un bois de Banksia, on peut retarder de 5 ans la mortalité causée par la maladie[46]. Les phosphites ralentissent également de 5 ans la progression de la maladie dans le boisement. Un éventuel incendie n'a pas d'impact sur ce ralentissement[47]. Une étude de 2003 a prouvé qu'en rinçant le sol avec une solution contenant 0,5 mM/L d'acide benzoïque on pouvait réduire significativement la taille des lésions de P. cinnamomi[48]. Des recherches sur la pourriture racinaire dans l'Ouest de l'Australie ont identifié un nouveau pathogène, P. multivora, isolé sur des eucalyptus malades et sur B. attenuata en 2009[49].
Culture et impact culturel

Les inflorescences jaune vif réparties régulièrement sur toute la plante la rendent assez attractive, sa forme naine et buissonnante étant préférée en horticulture[50]. Toutes les formes de Banksia attenuata requièrent un bon drainage, un sol sableux et une exposition ensoleillée, avec un pH entre 5,5 et 7[8]. Ils sont sensibles à Phytophthora cinnamomi, et supportent mal les climats humides[18]. Les graines n'ont pas besoin de traitement particulier, et mettent 16 à 49 jours pour germer[51]. Les graines sont très sensibles aux maladies fongiques[18]. Il faut attendre 4 à 6 ans pour obtenir un arbuste à maturité, capable de fleurir, à partir d'une graine[8]. Diverses tentatives pour améliorer leur adaptabilité aux climats humides comme le greffage ont échoué[18].
Les inflorescences sont utilisées par l'industrie de la fleur coupée[8], notamment en Australie-Occidentale[50].
Les Aborigènes d'Australie plaçaient les inflorescences dans un trou recouvert de feuilles de Melaleuca et rempli d'eau afin d'en faire une boisson. Cette espèce a pu, avec B. aemula, inspirer May Gibbs pour créer le personnage antagoniste qui a la forme d'une inflorescence de Banksia dans le livre pour enfants Snugglepot and Cuddlepie. Cette espèce était familière à Gibbs, qui l'avait côtoyée dans son enfance, même si c'est la seconde espèce citée qui correspond le plus aux descriptions[8]. L'artiste Marianne North a peint une toile assez populaire représentant un Banksia attenuata lors de son séjour en Australie entre 1880 et 1881[8] - [52].
Notes et références
- (en) Richard M Cowling et Byron B. Lamont, « Variation in serotiny of three Banksia species along a climatic gradient », Australian Journal of Ecology, vol. 10, no 3, , p. 345–50 (DOI 10.1111/j.1442-9993.1985.tb00895.x)
- (en) Alex S. George, « The Genus Banksia L.f. (Proteaceae) », Nuytsia, vol. 3, no 3, , p. 239–473 (ISSN 0085-4417)
- (en) Alex S. George, The Banksia Book, Kenthurst, New South Wales, Kangaroo Press, , 3e éd. (ISBN 0-86417-818-2), p. 133
- (en) George, Alex S. (dir.), Flora of Australia, vol. 17B, CSIRO Publishing / Australian Biological Resources Study, , 175–251 p. (ISBN 0-643-06454-0)
- The Banksia Atlas, p. 54–55
- (en) Richard M. Cowling et Byron B. Lamont, « Seed bank dynamics in four co-occurring Banksia species », Journal of Ecology, vol. 75, no 2, , p. 289–302 (lire en ligne)
- (en) Sue J. Wooller et Ronald D Wooller, « Seed set in two sympatric banksias, Banksia attenuata and B. baxteri », Australian Journal of Botany, vol. 49, , p. 597–602
- (en) Kevin Collins, Kathy Collins et Alex S. George, Banksias, Melbourne, Victoria, Bloomings Books, , 68, 150–51 (ISBN 978-1-876473-68-6 et 1-876473-68-1)
- (en) W.D. Stock, J.S. Pate et E. Rasins, « Seed developmental patterns in Banksia attenuata R.Br. and B. laricina C. Gardner in relation to mechanical defence costs », New Phytologist, vol. 117, , p. 109–14 (DOI 10.1111/j.1469-8137.1991.tb00950.x)
- « 'Banksia cylindrostachya Lindl. », Australian Plant Name Index (APNI), IBIS database, Centre for Plant Biodiversity Research, Australian Government
- « 'Sirmuellera attenuata (R.Br.) Kuntze », Australian Plant Name Index (APNI), IBIS database, Centre for Plant Biodiversity Research, Australian Government
- (en) Carl Meissner, « Proteaceae », Prodromus Systematis Naturalis Regni Vegetabilis, Paris, A. P. de Candolle, vol. 14,
- (en) George Bentham, « Banksia », Flora Australiensis, Londres, L. Reeve & Co, vol. 5, , p. 541–62
- (en) Kevin Thiele et Pauline Y. Ladiges, « A cladistic analysis of Banksia (Proteaceae) », Australian Systematic Botany (en), vol. 9, no 5, , p. 661–733 (DOI 10.1071/SB9960661)
- (en) Austin R. Mast, « Molecular systematics of subtribe Banksiinae (Banksia and Dryandra; Proteaceae) based on cpDNA and nrDNA sequence data: implications for taxonomy and biogeography », Australian Systematic Botany, vol. 11, nos 3–4, , p. 321–42 (DOI 10.1071/SB97026)
- (en) Austin R.Mast et Thomas J.Givnish, « Historical biogeography and the origin of stomatal distributions in Banksia and Dryandra (Proteaceae) based on their cpDNA phylogeny », American Journal of Botany, vol. 89, no 8, , p. 1311–23 (ISSN 0002-9122, DOI 10.3732/ajb.89.8.1311, lire en ligne)
- (en) Austin Mast, H Eric. et Shawn P. Havery, « An assessment of old and new DNA sequence evidence for the paraphyly of Banksia with respect to Dryandra (Proteaceae) », Australian Systematic Botany, CSIRO Publishing / Australian Systematic Botany Society, vol. 18, no 1, , p. 75–88 (DOI 10.1071/SB04015)
- (en) John Wrigley et Murray Fagg, Banksias, Waratahs and Grevilleas, Sydney, Angus & Robertson, , 17, 88 (ISBN 0-207-17277-3)
- (en) Austin R. Mast et Kevin Thiele, « The transfer of Dryandra R.Br. to Banksia L.f. (Proteaceae) », Australian Systematic Botany, vol. 20, no 1, , p. 63–71 (DOI 10.1071/SB06016)
- (en) Byron B. Lamont et Adrienne Markey, « Biogeography of fire-killed and resprouting Banksia species in South-western Australia », Australian Journal of Botany, vol. 43, no 3, , p. 283–303 (DOI 10.1071/BT9950283)
- (en) N.J. Enright et B.B. Lamont, « Seed banks, fire season, safe sites and seedling recruitment in five co-occurring Banksia species », Journal of Ecology, vol. 77, no 4, , p. 1111–22 (lire en ligne)
- (en) Richard M. Cowling et Byron B. Lamont, « Seed release in Banksia: the role of wet-dry cycles », Australian Journal of Ecology, vol. 10, no 2, , p. 169–71 (DOI 10.1111/j.1442-9993.1985.tb00878.x)
- (en) Richard M. Cowling et Byron B. Lamont, « Post-fire recruitment of four co-occurring Banksia species », Journal of Applied Ecology, vol. 24, no 2, , p. 645–58 (lire en ligne)
- (en) N. J. Enright et B. B. Lamont, « Recruitment variability in the resprouting shrub Banksia attenuata and non-sprouting congeners in the northern sandplain heaths of southwestern Australia », Acta Oecologica-International Journal of Ecology, vol. 13, no 6, , p. 727–41
- (en) J. Groeneveld, N.J. Enright, B.B. Lamont et C. Wissel, « A spatial model of coexistence among three Banksia species along a topographic gradient in fire-prone shrublands », Journal of Ecology, vol. 90, no 5, , p. 762–74 (DOI 10.1046/j.1365-2745.2002.00712.x, lire en ligne)
- (en) Tianhua He, Byron B. Lamont, Siegfried L. Krauss, Neal J.Enright et Ben P. Miller, « Long-distance dispersal of seeds in the fire-tolerant shrub Banksia attenuata », Ecography, vol. 32, no 4, , p. 571–80 (DOI 10.1111/j.1600-0587.2008.05689.x)
- (en) John K. Scott, « Estimation of the outcrossing oate for Banksia attenuata R. Br. and Banksia menziesii R. Br. (Proteaceae) », Australian Journal of Botany, vol. 28, no 1, , p. 53–59
- (en) « Specimen Report », sur Museum Victoria website: Bioinformatics, Melbourne, Australie, Museum Victoria, (consulté le )
- (en) R.A Tassone et J.D. Maier, « Abundance of Arthropods in Tree Canopies of Banksia Woodland on the Swan Coastal Plain », Journal of the Royal Society of Western Australia, vol. 80, , p. 281–86
- R.D. Barker et W.J.M. Vestjens, The Food of Australian Birds, Melbourne University Press, , 1:331, 2:458 (ISBN 0-643-05006-X)
- (en) John K. Scott et Robert Black, « Selective predation by White-tailed Black Cockatoos on fruit of Banksia attenuata containing the seed-eating weevil Alphitopis nivea », Australian Wildlife Research, vol. 8, no 2, , p. 421–30 (DOI 10.1071/WR9810421)
- (en) Delbert Weins, Marilyn Renfree et Ronald D. Wooller, « Pollen loads of Honey possums (Tarsipes spencerae) and non-flying mammal pollination in South-western Australia », Annals of the Missouri Botanic Gardens, vol. 66, , p. 830–38 (lire en ligne, consulté le )
- (en) Petroc Sumner, Catherine A. Arrese et Julian C. Partridge, « The ecology of visual pigment tuning in an Australian marsupial: the honey possum Tarsipes rostratus », Journal of Experimental Biology, vol. 208, no 10, , p. 1803–15 (DOI 10.1242/jeb.01610, lire en ligne)
- (en) C.C. Jordan, M.H. Brims, E.J. Speijers et E.M. Davison, « Myxomycetes on the bark of Banksia attenuata and B. menziesii (Proteaceae) », Australian Journal of Botany, vol. 54, no 4, , p. 357–65 (DOI 10.1071/BT05079)
- (en) Noel C. Beadle, « Some aspects of the ecology and physiology of Australian xeromorphic plants », Australian Journal of Science, vol. 30, , p. 348–355
- (en) A.B. Low et Byron B. Lamont, « Aerial and below-ground phytomass of Banksia Scrub-heath at Eneabba, South-western Australia », Australian Journal of Botany, vol. 38, no 4, , p. 351–59
- (en) Petra Marschner, Pauline F. Grierson et Zed Rengel, « Microbial community composition and functioning in the rhizosphere of three Banksia species in native woodland in Western Australia », Applied Soil Ecology, vol. 28, no 3, , p. 191–201 (DOI 10.1071/BT05081)
- (en) Byron B. Lamont et Stephen M. Bergl, « Water relations, shoot and root architecture, and phenology of three co-occurring Banksia species: no evidence for niche differentiation in the pattern of water use », Oikos, vol. 60, no 3, , p. 291–98 (lire en ligne)
- (en) R.H. Froend et P.L. Drake, « Defining phreatophyte response to reduced water availability: preliminary investigations on the use of xylem cavitation vulnerability in Banksia woodland species », Australian Journal of Botany, vol. 54, no 2, , p. 173–79 (DOI 10.1071/BT05081)
- (en) N.J. Enright et Byron B. Lamont, « Survival, growth and water relations of Banksia seedlings on a sand mine rehabilitation site and adjacent scrub-heath sites », Journal of Applied Ecology, vol. 29, no 3, , p. 663–71 (lire en ligne)
- (en) Judith L. Fisher, Erik J.Veneklaas, Hans Lambers et William A. Loneragan, « Enhanced soil and leaf nutrient status of a Western Australian Banksia woodland community invaded by Ehrharta calycina and Pelargonium capitatum », Plant and Soil, vol. 284, nos 1–2, , p. 253–64 (lire en ligne)
- (en) T.A. McCredie, K.W. Dixon et K. Sivasithamparam, « Variability in the resistance of Banksia L.f. species to Phytophthora cinnamomi Rands », Australian Journal of Botany, vol. 33, no 6, , p. 629–37 (DOI 10.1071/BT9850629)
- (en)[PDF] « Part 2, Appendix 4: The responses of native Australian plant species to Phytophthora cinnamomi », sur Management of Phytophthora cinnamomi for Biodiversity Conservation in Australia, Department of the Environment and Heritage, Australian Government, (consulté le )
- (en) T.C.J. Hill, J.T. Tippett et B.L. Shearer, « Invasion of Bassendean dune Banksia woodland by Phytophthora cinnamomi », Australian Journal of Botany, vol. 42, no 6, , p. 725–38 (DOI 10.1071/BT9940725)
- (en) C.L. Bishop, G.W. Wardell-Johnson et M.R. Williams, « Community-level changes in Banksia woodland following plant pathogen invasion in the Southwest Australian Floristic Region », Journal of Vegetation Science, vol. 21, no 5, , p. 888–98 (DOI 10.1111/j.1654-1103.2010.01194.x)
- (en) Bryan L. Shearer et Richard G. Fairman, « A stem injection of phosphite protects Banksia species and Eucalyptus marginata from Phytophthora cinnamomi for at least four years », Australasian Plant Pathology, vol. 36, no 1, , p. 78–86 (DOI 10.1071/AP06085)
- (en) Bryan L. Shearer et Richard G. Fairman, « Phosphite reduces disease extension of a Phytophthora cinnamomi front in Banksia woodland, even after fire », Australasian Plant Pathology, vol. 33, no 2, , p. 249–54 (DOI 10.1071/AP04002)
- (en) Mia Williams, Tissa Senaratna, Kingsley Dixon et Krishnapillai Sivasithamparam, « Benzoic acid induces tolerance to biotic stress caused by Phytophthora cinnamomi in Banksia attenuata », Plant Growth Regulation, vol. 41, , p. 89–91 (lire en ligne)
- (en) P.M. Scott, T.I. Burgess, P.A. Barber, B.L. Shearer, M.J.C. Stukely, G.E.St.J. Hardy et T. Jung, « Phytophthora multivora sp nov., a new species recovered from declining Eucalyptus, Banksia, Agonis and other plant species in Western Australia », Persoonia, vol. 22, , p. 1–13 (PMCID 2789538)
- (en) Lewis J. Matthews, The Protea Book : A Guide to Cultivated Proteaceae, Portland, Oregon, Timber Press, , 184 p. (ISBN 0-88192-553-5, lire en ligne), p. 39
- (en) Luke Sweedman et David Merritt, Australian Seeds : A Guide to their Collection, Identification and Biology, CSIRO Publishing, (ISBN 0-643-09298-6), p. 202
- (en)« A West Australian Banksia », sur Kew Gardens website, Richmond, Surrey, Board of Trustees of the Royal Botanic Gardens, Kew (consulté le )
Liens externes
- Ressources relatives au vivant :
- Global Biodiversity Information Facility
- (en) Australian Plant Name Index
- (en) Flora of Australia Online
- (en) FloraBase
- (en) Germplasm Resources Information Network
- (mul + en) iNaturalist
- (en) International Plant Names Index
- (en) The Plant List
- (en) Plants of the World Online
- (en) Tropicos
- (en) Union internationale pour la conservation de la nature
- (en) Référence FloraBase (Australie-Occidentale) : classification Banksia attenuata (+ photos + répartition + description) (consulté le )
- (en) Référence NCBI : Banksia attenuata (taxons inclus) (consulté le )
- (en) Référence GRIN : espèce Banksia attenuata R. Br. (consulté le )
- (en) Référence uBio : site déclaré ici indisponible le 7 avril 2023