Équilibre ponctué
La théorie des équilibres ponctués, aussi appelée ponctualisme ou ponctuationnisme, est un développement de la théorie de l'évolution proposée par deux paléontologues américains, Stephen Jay Gould et Niles Eldredge. Elle postule que l'évolution comprend de longues périodes d'équilibre, ou quasi-équilibre, ponctuées de brèves périodes de changements importants comme la spéciation ou les extinctions. Elle décrit l'évolution de la vie sur Terre sur un modèle accordant le darwinisme avec les hiatus fossilifères et avec les traces de grands bouleversements environnementaux passés que la théorie gradualiste (en) n'expliquait pas.
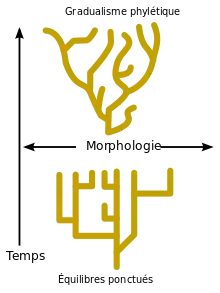
Apparition et contenu de la théorie
La théorie des équilibres ponctués, dont Pierre Trémaux fut le précurseur au XIXe siècle, a été présentée par Niles Eldredge et Stephen Jay Gould en 1972, dans un article intitulé : Punctuated equilibrium : an alternative to phyletic gradualism dans l'ouvrage collectif Models in Paleobiology. La théorie fut initialement très critiquée mais finit par emporter l'adhésion d'une majorité de paléontologues.
Gould et Eldredge ont présenté leur théorie comme une alternative au « gradualisme phylétique (en) », l'un des postulats de la théorie de l'évolution alors en vigueur (connue sous le nom de « Théorie synthétique de l'évolution » et issue du néo-darwinisme). Selon cette théorie, l'évolution morphologique des espèces se produirait par modifications très lentes et discontinues d'une même population au cours du temps par le jeu des mutations et de la sélection naturelle.
Toutefois, il semble que, bien avant Niles Eldredge et Stephen Jay Gould, Charles Darwin lui-même puisse être considéré comme un précurseur de la théorie des équilibres ponctués. Selon Patrick Tort [1], la lecture attentive de l'Origine des espèces « …paraît relativiser beaucoup la nouveauté revendiquée par le modèle des équilibres ponctués. »
La criticalité auto-organisée (Self-organized criticality ou SOC), qui montre comment la perturbation génère de nouvelles complexités, c'est-à-dire de la diversité et de nouvelles adaptations, s'applique à l'équilibre ponctué de Gould.
Fondement : vitesse de spéciation
Dérive génétique et sélection naturelle
Le phénomène de mutation génétique crée constamment de nouveaux allèles, entretenant ainsi une variabilité dans la population. Ces différents allèles peuvent réaliser au niveau individuel des phénotypes légèrement différents (ou au contraire être relativement neutres dans leur expression). La dérive génétique est à la fois permanente et aveugle, et donc tend à produire une dispersion toujours croissante sur les phénotypes de cette population.
Les variations individuelles des phénotypes peuvent entraîner des différences dans la sélection naturelle et le succès reproductif des individus. La sélection naturelle conduit donc, non pas à assurer « la survie des plus aptes », mais plus exactement à assurer l'élimination des moins aptes. Dans ce cas, les allèles statistiquement associés à un moindre succès reproductif (pour une raison ou une autre) seront sélectionnés négativement, et ceux au contraire qui confèrent statistiquement un avantage reproductif verront leur fréquence augmenter dans le pool génétique. Cet élagage continuel tend donc à s'opposer à la dérive génétique, et à réduire la variabilité de la population.
Le phénotype d'une population est donc le fruit d'un équilibre entre deux processus antagonistes : la dérive génétique, qui tend à augmenter la dispersion, et la sélection naturelle, qui élimine les versions sub-optimales.
Stabilité des phénotypes
Si une certaine direction de dérive du pool génétique permet une meilleure adaptation, l'élagage se fera au détriment des autres directions, et la population évoluera dans le sens d'une meilleure adaptation. Le phénotype d'une espèce tend donc, en pratique, vers celui qui assure en moyenne le meilleur avantage sélectif aux individus sur l'ensemble de son aire de répartition (aire sur laquelle est assuré le brassage génétique du pool génétique).
Mais une fois atteint un optimum local, un sommet d'adaptation, aucune dérive du phénotype n'est plus performante que cet optimum. L'effet de la sélection naturelle est alors de stabiliser le pool génétique vers un contenu qui, dans les générations passées, a été statistiquement optimal pour les individus recevant le phénotype associé, puisqu'assurant le meilleur succès reproductif possible.
Aussi longtemps que l'écosystème dans lequel se place une espèce est stable, son phénotype sera donc stable, puisqu'il ne peut pas s'écarter significativement de son optimum. L'espèce paraît stable, non pas qu'il n'y a pas de dérive génétique, mais parce toute évolution serait désavantageuse. De ce fait, il est exceptionnel de pouvoir observer une spéciation en cours.
Vitesse évolutive
Ce phénotype reste donc stable tant que les conditions écologiques le sont. Inversement, si les conditions écologiques d'une espèce varient, le pool génétique subira à nouveau une pression de sélection orientée, qui fera évoluer le phénotype.
L'évolution associée à un avantage reproductif peut être très rapide à l'échelle des temps géologiques. Ainsi, si les conditions écologiques font qu'une petite taille est légèrement avantagée par rapport aux grandes, si bien qu'à chaque génération la taille moyenne de la population diminue de un pour mille (ce qui représenterait deux millimètres à l'échelle humaine), il suffit de 700 générations pour que la taille moyenne soit divisée par deux (). S'il s'agit de générations humaines, cette évolution drastique se fera en à peine 20 000 ans.

Par exemple, certaines nouvelles espèces de souris comme celles de Madère n'ont nécessité qu'entre 500 et 1 000 ans pour apparaître (en fonction de l’origine viking ou portugaise qu’on leur attribue) ce qui représente environ 2 000 à 4 000 générations soit une vitesse extrêmement rapide.
De toute évidence, il peut exister des situations où les conditions écologiques évoluent lentement : la dérive du sous-continent Indien, par exemple, en a changé continument les conditions climatiques, conduisant les espèces endémiques à une adaptation tout aussi continue. On est alors dans un cas de « gradualisme phylogénétique », non pas parce que l'évolution des espèces est intrinsèquement lente, mais parce que le point d'optimum du pool génétique se déplace lentement avec l'évolution de l'environnement écologique.
Mais le changement des conditions écologiques peut plus vraisemblablement prendre la forme d'une catastrophe : suppression d'une barrière, introduction d'un nouveau prédateur, création d'une discontinuité dans l'aire de répartition, etc. Dans ce cas, l'évolution peut au contraire être très rapide, et ne pas laisser de trace dans les séries paléographiques.
Cas d'évolution rapide : Podarcis siculus

En 2004, une équipe scientifique dirigée par Duncan Irschick et Anthony Herrel s'est rendue sur l'île de Hrid Podmrčaru en mer Adriatique afin d'observer une espèce de lézard connu en France sous le nom de lézard des ruines. Ce lézard fut introduit sur l'île en 1971 par l'équipe du professeur Eviatar Nevo, un spécialiste de l'évolution, qui n'avait pu poursuivre l'observation de ce projet en raison des conflits yougoslaves. 36 années plus tard, Duncan Irschick et Anthony Herrel observent des changements notables dans la colonie de Podarcis siculus présente sur l'île. Les analyses d'ADN mitochondriales ont montré qu'il s'agit toujours de la même espèce Podarcis siculus, mais que des différences morphologiques notables tendent à distinguer cette population. Les spécimens étudiés sont plus grands que leur congénères du continent, leur mâchoire, plus puissante. Leur régime alimentaire s'est également modifié, essentiellement insectivores à l'origine, ils sont devenus majoritairement herbivores, et des valves cæcales sont apparues au niveau des intestins permettant de faciliter la digestion des herbes. Cette découverte pourrait corroborer le modèle d'évolution rapide développé dans la théorie des équilibres ponctués[2].
Critiques de la théorie
Débat « catastrophisme » – « gradualisme »
Le catastrophisme a précédé le gradualisme : les récits bibliques sont catastrophistes (Déluge, Apocalypse) et donc les créationnistes sont catastrophistes. Les premiers paléontologues tels le Français Georges Cuvier, le furent aussi (toutefois Cuvier croyait non pas à un, mais à plusieurs déluges). Dans un contexte de lutte idéologique entre les partisans d'une interprétation littérale des livres religieux (c'est-à-dire de l'intrusion de la religion dans le champ scientifique) et les chercheurs partisans d'une indépendance scientifique par rapport au champ religieux, le catastrophisme fut combattu par les théories gradualistes pour lesquelles l'évolution, aussi bien géologique (Charles Lyell, Richard Owen) que biologique (Charles Darwin, Ernst Mayr) n'agissait que très lentement sur d'immenses durées, notre monde actuel n'étant que le produit de l'accumulation de tout petits changements graduels. Le gradualisme expliquait que si les fossiles et les couches géologiques ont des « blancs », ce n'est pas parce que les changements d'ères furent violents et rapides, mais simplement parce que le grand livre de la Terre avait perdu des pages (pertes dues à la rareté de la fossilisation, au manque de dépôts sédimentaires ou à l'érosion). C'étaient les fameux « chaînons manquants ».
Mais les partisans de la théorie des équilibres ponctués affirment que ces manques sont très marqués dans les archives géologiques : la plupart d'entre eux ne peuvent pas s'expliquer de façon satisfaisante uniquement par la rareté de la fossilisation, le manque de dépôts et l'érosion, et le gradualisme ne pouvait expliquer ni les changements brutaux de faunes d'une couche géologique à l'autre, ni les changements tout aussi violents du climat, inscrits dans ces mêmes couches. Selon les partisans de la théorie des équilibres ponctués, le gradualisme est en contradiction avec le contenu des archives fossiles : selon eux, celles-ci ne contiendraient en fait que rarement les formes intermédiaires unissant une espèce à l'autre. En réalité, les espèces se caractériseraient à la fois par une apparition abrupte dans le registre fossile (ce qui peut signifier qu'elle s'est étendue sur quelques milliers d'années, un temps géologiquement court par rapport aux millions d'années postulés par le gradualisme phylétique) et par une grande stabilité à la suite de leur apparition. Cette période de stagnation morphologique des espèces (s'accompagnant tout au plus de quelques modifications mineures et réversibles) est appelée « stase » par les ponctualistes. Gould ira jusqu'à écrire que « l'extrême rareté des formes intermédiaires est le secret professionnel de la paléontologie ».
Selon Gould et Eldredge, la théorie des équilibres ponctués ne fait que résoudre une contradiction inhérente à la Théorie Synthétique : en effet, le célèbre biologiste Ernst Mayr, qui avait forgé cette théorie, avait soutenu un modèle de spéciation (c'est-à-dire de formation de nouvelles espèces) selon lequel les nouvelles espèces apparaissent quand une petite population d'une espèce donnée se trouve isolée de son aire de répartition d'origine (et donc de la population « mère »). Celle-ci connaîtrait alors une série rapide de changements morphologiques et génétiques avant de devenir une nouvelle espèce à part entière : c'est le modèle de spéciation péripatrique[3]. Les ponctualistes soutiennent que la théorie des équilibres ponctués ne fait qu'appliquer aux organismes fossiles ce modèle, tiré par les biologistes de l'observation des organismes modernes : les espèces se forment rapidement (voir plus bas l'évolution fulgurante, en 36 ans, du lézard insulaire Podarcis siculus), à partir de petites populations isolées (ce qui explique la rareté des intermédiaires), puis stagnent morphologiquement une fois établies. Ainsi s'expliquerait l'observation des équilibres ponctués.
Toutefois des exemples de gradualisme phylétique existent et ont été documentés dans le registre fossile, notamment chez les campagnols et certains trilobites. On a reproché à Gould et à Eldredge d'avoir sous-estimé la fréquence de ces exemples ou de ne pas avoir pris en compte le fait que l'absence d'intermédiaires peut être plutôt due à un manque de recherches ou au caractère lacunaire des archives fossiles qu'à leur réelle rareté. On a aussi pu affirmer que les paléontologues nommaient arbitrairement des espèces qu'ils découpaient au sein d'un continuum d'organismes se transformant progressivement au long des temps géologiques, créant ainsi artificiellement des discontinuités ensuite récupérées comme des preuves par les ponctualistes.
Les ponctualistes ont répondu à la première objection par des études de fréquence relative couvrant un large nombre d'espèces, mais la conclusion de ces études a là aussi été variable suivant les cas, certaines étant favorables aux ponctualistes et d'autres aux gradualistes : il semblerait en fait que les deux modes d'évolution existent, selon les groupes, voire selon les caractères considérés. Ils ont également répondu à la troisième par des études évaluant de façon quantitative les modifications morphologiques observées dans les fossiles, sans prendre en compte la classification en espèces : ils ont ainsi pu montrer objectivement, au moins dans certains cas, la réalité de l'évolution par équilibres ponctués.
Importance de la théorie
Certains auteurs, tels que Richard Dawkins ont affirmé que, même si elle était exacte, elle n'avait pas le caractère révolutionnaire que lui prêtaient ses fondateurs. Selon Dawkins, la théorie des équilibres ponctués ne serait « qu'une vague mineure sur l'océan du néodarwinisme ». Stephen Jay Gould a répondu en détail à cette affirmation de Dawkins, Gould la décrivant comme étant mesquine et déplacée, car ignorant les implications de la théorie des équilibres ponctués en matière de macro-évolution[4].
Interprétations détournées de la théorie des équilibres ponctués
La théorie des équilibres ponctués a été à plusieurs reprises caricaturée ou mal comprise dans les concepts qu'elle développe.
Données négatives
Une objection fréquente est que la théorie des équilibres ponctués se fonde sur une absence de fossiles transitoires, mais une absence ne peut être scientifiquement qualifiée comme une preuve significative : la qualité des archives fossiles étant très aléatoire, de telles lacunes ne sont pas indicatives de l'évolution suivie par des espèces.
Gould souligne que la théorie des équilibres ponctués ne s'appuie pas uniquement sur des données négatives comme l'absence de fossiles transitoires, mais aussi sur des données positives, comme l'existence des périodes de stase, ces longues durées où une espèce ne connaît aucun changement morphologique majeur. Selon un aphorisme célèbre de Stephen Jay Gould, « stasis is data » (la stase est une donnée) : si les paléontologues ont souvent eu tendance à négliger d'étudier la simple absence de changement quand ils l'observaient, elle n'en constitue pas moins selon Gould une donnée réelle.
Formes intermédiaires
La théorie des équilibres ponctués est parfois interprétée comme affirmant qu'il n'existe pas, ou peu, de fossiles de formes intermédiaires, et se présentant comme une explication à cette absence.
Gould remarque que la théorie des équilibres ponctués concerne uniquement la formation de nouvelles espèces, donc l'absence de formes intermédiaires entre espèces différentes. L'espèce est l'un des échelons les plus bas de la classification du monde vivant : les espèces intermédiaires entre groupes plus vastes (par exemple entre reptiles et mammifères ou entre dinosaures et oiseaux) sont au contraire nombreuses. « Les formes intermédiaires manquent habituellement au niveau des espèces, mais elles abondent entre groupes plus vastes », écrit Gould.
Monstre prometteur
Une partie des objections formulées contre la théorie des équilibres ponctués la rapprochent d'une version actualisée de la théorie du monstre prometteur. Cette théorie de l'évolution fut initialement élaborée dans les années 1930 par le généticien Richard Goldschmidt, et suppose qu'une seule mutation de grande ampleur pourrait donner naissance à une nouvelle espèce, voire à un nouveau « grand groupe » d'êtres vivants en donnant naissance en une seule étape à un nouveau plan d'organisation.
La théorie des équilibres ponctués affirme que la formation de nouvelles espèces est instantanée géologiquement parlant, mais peut impliquer une durée réelle de quelques milliers d'années, et impliquer un nombre significatif de petites mutations plutôt qu'une seule mutation de grande ampleur. La théorie du monstre prometteur visait à expliquer la naissance de grands groupes caractérisés par des innovations évolutives radicales. La théorie des équilibres ponctués se focalise au contraire sur le phénomène de spéciation.
Le point commun entre ces deux théories est de supposer un changement évolutif rapide. Gould s'est parfois déclaré favorable à une forme modérée de la théorie du monstre prometteur, mais ces déclarations étaient, dans leur contexte, sans rapport direct avec ses travaux sur la théorie des équilibres ponctués.
Abus idéologiques
Des courants créationnistes protestants détournent les objections soulevées par les ponctualistes contre les imperfections du modèle développé dans la théorie synthétique de l'évolution, et s'en servent pour discréditer l'ensemble des théories de l'évolution découlant des hypothèses formulées par Darwin. Néanmoins la théorie des équilibres ponctuées et les travaux scientifiques qui y sont liés ne remettent pas en cause les fondements de la théorie de l'évolution de Darwin : des variations héréditaires aléatoires sélectionnées par le milieu où ils vivent.
Les ponctualistes et les partisans de l'uniformitarisme, tels Philippe Taquet ou Léonard Ginsburg, ne renient pas les principes de l'évolution et n'admettent pas la confusion entre démarche scientifique, qui relève de la recherche, de l'expérimentation et de la vérification continuelles, et la démarche religieuse, qui relève de la fidélité à une foi et à des rites. Tous soulignent que les preuves de l'évolution se sont à tel point accumulées, que celles-ci relèvent assurément d'une théorie, mais tout aussi fondée que celles des probabilités, des tenseurs ou de la gravitation, c'est-à-dire au sens scientifique d'édifice cohérent et non au sens commun d'hypothèse. Le Vatican lui-même a d'ailleurs admis officiellement qu'il y voyait « davantage qu'une hypothèse »[5].
Le concept de ponctuation au-delà des sciences naturelles
Des sciences naturelles aux sciences humaines
Le concept d'« équilibres ponctués » est parfois étendu à d'autres domaines que la Paléontologie et les sciences de l'évolution. François Terrasson considère par exemple que le principe peut s'appliquer à la Géonomie et aux sciences humaines. On peut aussi l'appliquer aux langues[6] et à l'évolution des mythes[7]. Dans cette perspective, les systèmes dynamiques étudiés peuvent être perçus comme ayant tendance à s’équilibrer et se stabiliser durant une période où les évolutions sont graduelles. Ces équilibres stabilisés peuvent parfois être ponctuellement bouleversés et s’effondrer brutalement[8].
L'humanité, une ponctuation ?
Selon Richard Leakey et Roger Lewin, la prolifération de l'espèce humaine et l'accroissement de ses capacités à modifier les environnements constituent une ponctuation, qui se traduira par une nouvelle phase d'extinction. Le débat est ouvert pour savoir si cette phase existe, si elle a déjà commencé, si les processus enclenchés sont ou non réversibles, si notre espèce y survivra.
Notes et références
- Note dans Patrick Tort, Darwin et le Darwinisme, Paris/58-Clamecy, PUF, coll. « Que sais-je ? », , 128 p. (ISBN 978-2-13-078769-3), p. 89.
- Voir site :(en) PNAS, vol. 105, n°12, pages 4792-4795 (25 mars 2008).
- Cédric Grimoult, L'évolution et les évolutionnistes de A à Z, Editions Ellipses, , p. 297
- Stephen Jay Gould, La Structure de la théorie de l'évolution, Éditions Gallimard, 2006, pages 1427-1430
- « Jean Paul II et l'évolution des espèces - Hominidés », sur hominides.com (consulté le ).
- (en) Quentin D. Atkinson, Andrew Meade, Chris Venditti, Simon J. Greenhill, Mark Pagel, Languages Evolve in Punctuational Bursts, Science, 319 (5863), 2008: 588.
- Julien d'Huy, Les mythes évolueraient par ponctuations, Mythologie française, 252, 2013, 8-12.
- « L’évolution : la naissance des espèces », dans Science et Vie, 173, décembre 1990; « L’histoire de la Vie », dans La Recherche, 296, mars 1997; « La valse des espèces », dans Pour la Science M 1930, juillet 2000 ; « Dans le secret des mondes disparus », dans Science et Vie, 213, décembre 2000; « L’évolution », dans La Recherche, 27, mai-juin 2007
Bibliographie
- Jules Carles, Les Origines de la Vie, PUF, coll. « Que sais-je ? », Paris, 1962
- André Cailleux, La Terre et son histoire, PUF, coll. « Que sais-je ? », Paris, 1978
- Richard Dawkins, L'Horloger aveugle, éditions Robert Laffont, 1989
- Vincent Courtillot, La Vie en catastrophes, Fayard, Paris, 1995
- Éric Buffetaut, Histoire de la paléontologie, PUF, coll. « Que sais-je ? », Paris, 1998
- Stephen Jay Gould, Le Pouce du panda, Grasset, 1982
- Stephen Jay Gould, Quand les poules auront des dents, Fayard, 1984
- Stephen Jay Gould, Aux racines du temps, Grasset, Paris, 1990
- Stephen Jay Gould, Le Livre de la Vie, Seuil, Paris, 1993
- (en) Niles Eldredge, The Pattern of Evolution, Freeman, 2000
- Stephen Jay Gould, La Structure de la théorie de l'évolution, Gallimard, 2006
- Jean-Claude Léonide, L'Évolution de l'amibe au cerveau humain en perspective élargie, Promothéa, Paris 1993
- (en) Michael Givel, Failure to Change through Multiple Policy Instruments and Venues the Tobacco Industry Policy Subsystem in the States from 1990 to 2003, University of Oklahoma, 2006
- Richard Leakey et Roger Lewin, La Sixième Extinction, Flammarion, Paris, 1995
- (en) Wilkins, J.S. and Nelson, G.J. (2008). Trémaux on species: A theory of allopatric speciation (and punctuated equilibrium) before Wagner. Archives of Philosophy of the Science. University of Pittsburgh USA. pdf: http://philsci-archive.pitt.edu/3881/1/Tremaux-on-species.pdf . Publié online comme format "pre-print" dans cette bibliothèque digitale et définitivement publié, exactement avec le même titre et texte, dans: History and Philosophy of the Life Sciences (Hist. Philos. Life Sci, 2008, 30:179-206).