Trophectoderme
Le trophectoderme est une couche de cellules externes de la morula puis du blastocyste.
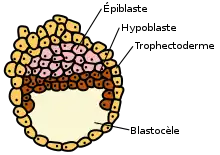
Lors de la première semaine du développement embryonnaire, l'embryon progresse de l'ampoule tubaire vers la cavité utérine. Durant ce trajet, il va subir de multiples modifications qui vont lui permettre d'aboutir au stade de blastocyste et au cours desquelles va apparaître le trophectoderme.
Formation du trophectoderme
La séparation physique et fonctionnelle de la masse cellulaire interne du trophectoderme est une caractéristique particulière du développement des mammifères et constitue la première spécification de la lignée cellulaire dans ces embryons. Après la fécondation dans l'oviducte, l'embryon de mammifère subit une série relativement lente de clivages pour produire une morula à huit cellules. Chaque cellule de la morula, augmente le contact de surface avec ses voisines dans un processus appelé compactage. Il en résulte une polarisation des cellules dans la morula et un clivage supplémentaire donnant un blastocyste d'environ 32 cellules[1]. Chez la souris, environ 12 cellules internes composent la nouvelle masse cellulaire interne et 20 à 24 cellules composent le trophectoderme environnant[2] - [3]. Il existe une variation entre les espèces de mammifères quant au nombre de cellules au compactage avec des embryons bovins montrant des différences liées au compactage dès 9-15 cellules et chez les lapins pas avant 32 cellules. Il existe également des variations entre embryons d'une même espèce[4].
Génétique
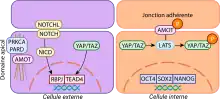
Cette compaction entraine la formation d'un domaine apicale pour les cellules situées à l'extérieur de la morula, on observe l'expression des éléments du cytosquelette kératine 8 et 18 qui interagissent avec le domaine apical. Ces éléments du cytosquelette ainsi que d'autres protéines membranaires vont participer à l'emprisonnement des protéines angiomotines. Ces protéines ne pouvant plus joué leurs rôles de régulation des jonctions adjacentes ou elles sont phosphorylé, la signalisation cellulaire étant interrompue les kinases Lats sont libérés entrainant l'activation de la voie de signalisation Hippo. Les kinases Lats phosphoryle à leurs tours les facteurs de transcription Yap et Taz qui grâce au domaine apical vont pouvoir entré dans le noyau de la cellule. Une fois dans le noyau et avec la coopération d'autres voies de signalisation cellulaire se lieront au facteur de transcription Tead4, activant à son tour un ensemble de gènes permettant la transformation des cellules totipotente en cellules du trophectoderme, ce facteur va également réprimer l'expression des gènes liés à la pluripotence tels que le facteur de transcription Sox2[5].
Devenir du trophectoderme
Souris et primates
Après l'apparition des cellules de l'hypoblaste et le passage au stade du blastocyste, le trophectoderme dit polaire qui est adjacent aux cellules de l'épiblaste va venir se fixer sur l'endomètre de l'utérus, c'est la Nidation. Les cellules du trophectoderme donneront les cellules du Cytotrophoblaste et Syncytiotrophoblaste qui seront à l'origine du placenta[5].
Autres mammifères
Chez les autres mammifères tels que les lapins, bovins et cochons. L'implantation de l'embryon sur les parois utérines est beaucoup plus tardive après la gastrulation. On observe alors une disparition des cellules du trophectoderme recouvrant les cellules de l'épiblaste, ces cellules du trophectoderme sont appelées membrane de Rauber. Le rôle de la disparition de ces cellules fait encore l'objet de recherche. Il a été montré chez les bovins que la disparition de ces cellules permettait d'empêcher une polarisation de l'embryon et d'aider à la formation de l'axe antéro-postérieur[5].
Références
- Lewis Wolpert, Principles of development, Oxford university press, (ISBN 978-0-19-927537-3, lire en ligne)
- Yusuke Marikawa et Vernadeth B. Alarcón, « Establishment of trophectoderm and inner cell mass lineages in the mouse embryo », Molecular Reproduction and Development, vol. 76, no 11, , p. 1019–1032 (ISSN 1098-2795, PMID 19479991, PMCID 2874917, DOI 10.1002/mrd.21057, lire en ligne, consulté le )
- Aneta Suwińska, Renata Czołowska, Wacław Ozdzeński et Andrzej K. Tarkowski, « Blastomeres of the mouse embryo lose totipotency after the fifth cleavage division: expression of Cdx2 and Oct4 and developmental potential of inner and outer blastomeres of 16- and 32-cell embryos », Developmental Biology, vol. 322, no 1, , p. 133–144 (ISSN 1095-564X, PMID 18692038, DOI 10.1016/j.ydbio.2008.07.019, lire en ligne, consulté le )
- Ewart W. Kuijk, Leonie du Puy, Helena TA van Tol et Henk P. Haagsman, « Validation of reference genes for quantitative RT-PCR studies in porcine oocytes and preimplantation embryos », BMC Developmental Biology, vol. 7, no 1, , p. 58 (ISSN 1471-213X, PMID 17540017, PMCID PMC1896162, DOI 10.1186/1471-213X-7-58, lire en ligne, consulté le )
- Wilhelm Bouchereau, « Déchiffrer le continuum de pluripotence dans l’embryon pré-implantatoire de lapin », {{Article}} : paramètre «
périodique» manquant, Lyon, (lire en ligne, consulté le )