Hypoblaste
L'hypoblaste (ou endoderme primitif) est l'une des deux couches distinctes issues de la masse cellulaire interne du blastocyste de mammifère, ou du blastodisque chez les reptiles et les oiseaux. L'hypoblaste forme le sac vitellin, qui à son tour donne naissance au chorion.
L'hypoblaste est une couche de cellules dans les embryons de poissons et d'amniotes. L'hypoblaste aide à déterminer les axes corporels de l'embryon et sa migration détermine les mouvements cellulaires qui accompagnent la formation de la ligne primitive et aide à l'orientation de l'embryon et à la création d'une symétrie bilatérale.
L'autre couche de la masse cellulaire interne, l'épiblaste, se différencie en trois feuillets embryonnaires primaires, l'ectoderme, le mésoderme et l'endoderme .
La structure
L'hypoblaste se trouve sous l'épiblaste et se compose de petites cellules cubiques[1]. L'hypoblaste chez les poissons (mais pas chez les oiseaux et les mammifères) contient les précurseurs de l'endoderme et du mésoderme[2]. Chez les oiseaux et les mammifères, il contient des précurseurs de l'endoderme extra-embryonnaire du sac vitellin [2].
Pour les mammifères
Origine
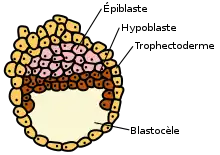
Dans l'embryogenèse des mammifères, la différenciation et la ségrégation des cellules dans la masse cellulaire interne du blastocyste produisent deux couches différentes - l'épiblaste et l'hypoblaste également appelé endoderme primitif[2].
Pour la souris, aux stades blastocytaire les cellules de la masse cellulaire interne co-expriment le gène marqueur de l'épiblaste pluripotent nanog et le gène différentiateur de l'hypoblaste gata6. Progressivement l'expression de ces gènes devient exclusive à certaines cellules donnant une répartition « poivre-et-sel » de cellules exprimant uniquement Gata6 ou Nanog, qui seront les précurseurs respectivement de l'hypoblaste et de l’épiblaste. Les cellules de l'hypoblaste vont ensuite migrer vers l'intérieur du blastocyste et progressivement recouvrir la face viscérale de l'épiblaste grâce à des mouvements cellulaires et des changements de polarité, les cellules qui ne se sont pas correctement différenciées dans un des deux types cellulaires ou mal positionnés sont éliminées par apoptose.
Pour l'Homme La formation d'hypoblastes dépend de la signalisation du facteur de croissance des fibroblastes , car la nouvelle population d'hypoblastes maintient l'expression de Gata6 en réponse au facteur de croissance des fibroblastes sécrété par l'épiblaste[3].
À la fin du stade blastocyste, les cellules de la masse cellulaire interne du rongeur se sont différenciées en lignées d'épiblastes et d'hypoblastes à la suite de migrations cellulaires et d'apoptoses. Elles perdent alors progressivement leur plasticité phénotypique et s'engagent de manière irréversible dans l'une ou l'autre lignée. Les cellules hypoblastiques adjacentes au sac vitellin se polarisent et forment un épithélium. L'épithélialisation constitue la dernière étape de la spécification des hypoblastes chez les rongeurs et les primates, qui sont maintenant prêts pour l'implantation[3].
Génétique
Pour rompre la présence initiale des deux gènes Gata6 et Nanog dans les cellules de la masse cellulaire interne, l'activation de la voie de signalisation des facteurs de croissance des fibroblastes dans les cellules qui deviendront l'hypoblaste. Pour la souris l'utilisation du gène Fgf4 induit la formation de cellules de l'hypoblaste proportionnellement à la dose utilisée. Fgf4 est un ligand qui est produit spécifiquement par les cellules de la masse cellulaire interne et de l'épiblaste. En l'absence du gène Nanog, Fgf4 provoque l'expression des gènes Sox17 et Gata4 dans les cellules présentant Gata6. Il a également été observé que les cellules dont la présence de Fgf4 est stoppée perdent progressivement l'expression du gène Gata6, de même que le blocage des récepteurs aux facteurs de croissance des fibroblastes provoque la disparition des cellules possédant Gata6 dans la masse cellulaire interne[4].
Fonction
Bien que l'hypoblaste ne contribue pas à l'embryon, il influence l'orientation de l'embryon[2]. L'hypoblaste contrôle également la formation de la ligne primitive[5]. L'absence d'hypoblastes entraîne la formation de multiples lignes primitives chez les embryons de poulet[6]. Le sac vitellin formé à l'aide de l'hypoblaste assure la bonne organogenèse du fœtus et l'échange de nutriments, de gaz et de déchets. Les cellules hypoblastes fournissent également des signaux chimiques qui spécifient la migration des cellules de l'épiblaste[2].
Pour les poissons
Chez les poissons, l'hypoblaste est la couche interne de la marge épaissie du blastoderme épibolisant dans l'embryon de poisson gastrulant[2].
Références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Hypoblast » (voir la liste des auteurs).
- « The Developing Human - 11th Edition », sur www.elsevier.com (consulté le )
- Michael Barresi et Scott Gilbert, Developmental Biology, 12th, (ISBN 978-1605358222)
- (en) Connor Ross et Thorsten E. Boroviak, « Origin and function of the yolk sac in primate embryogenesis », Nature Communications, vol. 11, no 1, , p. 3760 (ISSN 2041-1723, DOI 10.1038/s41467-020-17575-w, lire en ligne, consulté le )
- Wilhelm Bouchereau, « Déchiffrer le continuum de pluripotence dans l’embryon pré-implantatoire de lapin », Université de Lyon (thèse), (lire en ligne, consulté le )
- Joaquim Egea, Christian Erlacher, Eloi Montanez et Ingo Burtscher, « Genetic ablation of FLRT3 reveals a novel morphogenetic function for the anterior visceral endoderm in suppressing mesoderm differentiation », Genes & Development, vol. 22, no 23, , p. 3349–3362 (ISSN 0890-9369, PMID 19056886, PMCID 2600762, DOI 10.1101/gad.486708, lire en ligne, consulté le )
- « Nodal antagonists in the anterior visceral endoderm prevent the formation of multiple primitive streaks », Dev Cell, vol. 3, no 5, , p. 745–56 (PMID 12431380, DOI 10.1016/S1534-5807(02)00321-0)