Système de sécrétion de type IV
Le système de sécrétion de type IV (aussi connu sous le nom T4SS) est un complexe protéique de sécrétion que l’on retrouve souvent chez les bactéries.
Il s’agit d’un moyen pour permettre le transport de protéines ou d’ADN à travers la membrane cellulaire.
Le T4SS est un système utilisé pour la conjugaison bactérienne, pour l’échange de matériel génétique (par exemple de l’ADN plasmidique) avec le milieu extra-cellulaire et pour délivrer des protéines dans des cellules cibles.
Les systèmes de sécrétion de type IV (SST4) sont divisés en quatre familles : IVA (regroupant les SST4-F et -P), IVB, IVGI et IVGP[1].
Le T4SS intervient par exemple pour l’insertion du plasmide dans les plantes avec Agrobacterium tumefaciens, la délivrance de toxine chez Bordetella pertussis et Legionella pneumophila ou encore avec le pilus sexuel F.
Fonction
Le système de sécrétion de type IV est un complexe protéique présent chez les cellules procaryotes utilisé pour le transport d’ADN, de protéines et d’autres molécules depuis le cytoplasme vers le milieu extra-cellulaire. Le T4SS est lié à la machinerie de conjugaison procaryote. Le système de sécrétion de type IV est très varié. Il est présent dans bactérie Gram négative, Gram positive et les Archaea.
Les systèmes de sécrétion de type IV sont regroupés sur la base de similitudes structurelles et génétiques. Ils sont ainsi souvent liés de manière assez lointaine.
- Les systèmes de types IVA sont similaires aux systèmes virB / D4 d’Agrobacterium tumefaciens.
- Les systèmes de types IVB sont similaires aux systèmes Dot/Icm trouvés dans les agents pathogènes intracellulaires tels que Legionella pneumophila.
- Les autres types ne ressemblent pas aux types IVA et IVB.
Les différents systèmes de sécrétion de type IV sont génétiquement différents et donc utilisent des protéines différentes. Cependant, les protéines entre les différents ensembles ont de fortes homologies ce qui les conduit à fonctionner de la même manière.
Les systèmes de sécrétion de type IV sont également classés par fonction en trois groupes principaux.
- Système conjugué : utilisés pour la transfert d’ADN via un contact cellule/cellule (conjugaison bactérienne).
- Système de libération et d’absorption d’ADN : utilisés pour échanger de l’ADN avec l’environnement extra-cellulaire.
- Système effecteur: utilisés pour transférer des protéines vers des cellules cibles.
Les systèmes de libération et d’absorption d’ADN jouent un rôle important dans le transfert de gènes, ce qui permet aux procaryotes de s’adapter à leur environnement (comme par exemple développer une résistance aux antibiotiques).
Les systèmes effecteurs permettent l’interaction avec des organismes plus gros. Ils interviennent pour administrer des toxines dans le cas de nombreux agents pathogènes pour l’homme. Ils interviennent dans des maladies comme les ulcères à l’estomac, la coqueluche ou la légionellose.
Structure
Actuellement, seule la structure de système de sécrétion de type IVA, qui sont présents dans les bactéries Gram négative, est bien décrite. Il est composé de 12 sous unités protéiques de VirB1 à VirB11 et de VirD4 dont les analogies existent dans tous les systèmes de type IVA. Les composants du système de sécrétion de type IV peuvent être séparés en trois groupes : l’échafaudage du canal de translocation, les ATPases et le pilus.
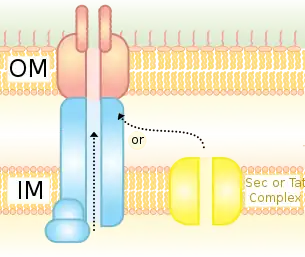
L’échafaudage du canal de translocation fait intervenir plusieurs aux protéines qui créent un canal entre l’espace extracellulaire et le cytoplasme à travers les membranes interne et externe de la bactérie. Ils contiennent VirB6 à VirB10. Le complexe central de l’échafaudage est composé de 14 copies de VirB7, VirB9 et VirB10 qui forment un canal cylindrique qui s’étend sur les deux membranes et relie le cytoplasme à l’espace extra-cellulaire.
Une seule protéine, VirB10 fait partie intégrante des membranes interne et externe. Il s’insère dans la membrane externe en utilisant une structure en tonneau hélicoïdal qui aide à former un canal entre les deux membranes. Il y a une ouverture sur l’extrémité cytoplasmique du canal, qui suivi d’une grande chambre et d’une seconde ouverture. La deuxième ouverture nécessite un changement de conformation pour permettre le passage du substrat du cytoplasme dans le canal. On pense que VirB6 ou VirB8 forment le pore de la membrane interne, car ce sont des protéines intégrales sur la membrane interne et ont un contact direct avec le substrat.
Les ATPases sont constitués de VirB4, VirB11 et VirD4 qui entraînent le mouvement du substrat vers le canal et fournissent de l’énergie au système de sécrétion de type quatre. VirB11 appartient à une classe de transporteur transmembranaire appelé « ATPases trafic ». VirB4 n’est pas très bien caractérisée.
Le pilus est composé de VirB2 et VirB5. VirB2 étant le composant principal. Chez A.tumefaciens, le pilus a un diamètre de 8 à 12nm et une longueur inférieure à 1 µm. Les pilis F, un autre type de pilus sont couramment étudiés. Ils sont beaucoup plus longs avec une longueur de 2 à 20 µm.
Mécanisme
En raison de la grande variété des systèmes de sécrétion de type quatre à la fois dans leur origine dans les fonctions, il est très difficile de formuler la mécanique du groupe dans son ensemble.
En général, une fois l’ADN emballé par le système de conjugaison, il est ensuite recruté par les ATPases et notamment la protéine de couplage VirD4. Il est ensuite transféré à travers le pilus. Chez A.tumefaciens, l’ADN traverse une chaîne enzymatique avant d’atteindre le pilus. L’ADN est ensuite recruté par VirD4 et VirB11, puis par les protéines transmembranaires (VirB6 et VirB8), déplacé vers VirB9, et enfin envoyé vers le Pilus (VirB2).
Références
- Gauthier Dangla-Pélissier, Nicolas Roux, Victoria Schmidt et Gaël Chambonnier, « The horizontal transfer of Pseudomonas aeruginosa PA14 ICE PAPI-1 is controlled by a transcriptional triad between TprA, NdpA2 and MvaT », Nucleic Acids Research, vol. 49, no 19, , p. 10956–10974 (ISSN 0305-1048 et 1362-4962, DOI 10.1093/nar/gkab827, lire en ligne, consulté le )