Pluteus (échinidé)
Pluteus (ou « echinopluteus ») est le nom qu'ont donné les scientifiques au premier stade larvaire des oursins, qui est encore planctonique.
_early_pluteus_width_ca._600_%C2%B5m_Helgoland_5.2006_9222.tif.jpg.webp)
Description
Le Pluteus est la première forme de l'oursin à la sortie de l’œuf, qui éclot en pleine mer. Ils sont totalement transparents et leur forme évoque une Tour Eiffel microscopique, pourvue de trois à six bras ciliés (chaque groupe d'oursins possède cependant des particularités dans la morphologie larvaire[1])[2]. Ces bras sont renforcés par un fin squelette calcaire, plus ou moins développé suivant les espèces, et facilement visible au microscope en lumière polarisée.
Les larves dérivent parmi le plancton pendant plusieurs semaines (parfois plusieurs mois, voire années[3]) où elles se nourrissent principalement de phytoplancton[4], puis se laissent couler vers le fond pour se fixer sur un substrat et y entamer leur métamorphose en petits oursins juvéniles, encore particulièrement vulnérables[5].
Ces larves pluteus ont une symétrie bilatérale, contrairement aux échinodermes adultes qui sont pentaradiés (symétrie radiale d'ordre 5) : ces larves sont donc la preuve que les échinodermes sont bien des bilatériens, la symétrie pentaradiaire n'étant acquise que secondairement, à la métamorphose[2].
Particularités et exceptions
Certaines espèces polaires ont développé un mode de reproduction différent, appelé « lecithotrophe » (la reproduction classique des oursins étant nommée « planctotrophe ») : la fécondation est toujours externe, mais se fait avec des gamètes plus gros et moins nombreux, dont le développement débouche directement sur de petits juvéniles, sans passer par le stade planctonique. Les femelles de ces espèces portent ainsi des poches d'incubation caractéristiques[4].
Des cas de reproduction asexuée ont été observés chez des larves de Dendraster excentricus, et seraient possibles chez d'autres espèces : dans des eaux riches en nutriments mais aussi en prédateurs, les larves planctoniques sont capables de se cloner par division, doublant ainsi leurs chances de survie[3].
Utilisation scientifique
L'oursin est un modèle très utilisé pour la recherche : réaliser une fécondation en laboratoire est relativement simple, et les élevages d'oursins sont faciles à maintenir et peu coûteux contrairement à de nombreux autres animaux modèles. De plus, leurs œufs et leurs larves sont transparents, ce qui permet une excellente visibilité des processus cellulaires. Les espèces les plus utilisées par la recherche scientifique sont notamment Arbacia punctulata et Strongylocentrotus purpuratus[6].
Les pluteus présentent par ailleurs des difformités si les concentrations de polluants dans l'eau dépassent un certain seuil [7] - [8] : ils sont donc utilisés en écotoxicologie comme indicateurs de pollution du milieu. De même, le pourcentage d'ovocytes fécondés diminue avec l'augmentation des polluants dans le milieu : la densité de larves est donc, elle aussi, signifiante de l'état de l'eau.
Pendant plus d'un siècle, les embryologistes ont utilisé A. punctulata comme modèle expérimental, car les nombreux embryons produits par une seule ponte se développent de manière synchrone[9] - [10].
En génétique, pendant des décennies, l'embryon d'oursin a été utilisé pour établir la théorie chromosomique de l'hérédité, la description des centrosomes, la parthénogenèse et la fécondation[11] - [12] - [13]. Les travaux de recherche au cours des trente dernières années, ont permis de comprendre des phénomènes tels que l'ARNm stable et le contrôle de la traduction génétique, l'isolement et la caractérisation du fuseau mitotique, et la réalisation que les principales protéines de structure de fuseau sont les microtubules[14] - [15].
Le premier échinoderme à avoir vu son génome entièrement séquencé fut l'oursin pourpre Strongylocentrotus purpuratus[16].
Galerie
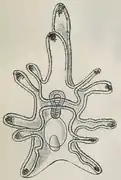 schéma de développement d'un pluteus.
schéma de développement d'un pluteus.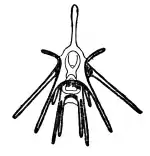 Autre schéma de pluteus.
Autre schéma de pluteus. Pluteus d'oursin noir dessiné par Ernst Haeckel.
Pluteus d'oursin noir dessiné par Ernst Haeckel.%252C_early_pluteus_ventral_view%252C_length_500_mu_Helgoland_5.JPEG.webp)
_early_pluteus_width_ca.JPEG.webp)
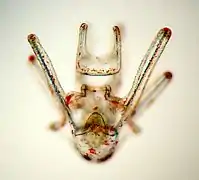
 Squelette calcaire d'un pluteus de C. subdepressus mis en évidence en lumière polarisée.
Squelette calcaire d'un pluteus de C. subdepressus mis en évidence en lumière polarisée.
Voir aussi
Liens internes
Les larves d'autres échinodermes :
- Ophiopluteus, larve d'Ophiuroidea (ophiure) ;
- Bipinnaria, Brachiolaria, larves d'Asteroidea (étoile de mer) ;
- Auricularia, larve d'Holothuroidea (concombre de mer).
Notes et références
- (en) « Introduction », sur l'Echinoid Directory du Natural History Museum.
- (en) « Larvae », sur l'Echinoid Directory du Natural History Museum.
- (en) Christopher Mah, « Cloning As Sand Dollar defense : Why run & hide when you can divide? », sur Echinoblog, .
- (en) « Reproduction and life history », sur l'Echinoid Directory du Natural History Museum.
- Alain Guille, Pierre Laboute et Jean-Louis Menou, Guide des étoiles de mer, oursins et autres échinodermes du lagon de Nouvelle-Calédonie, ORSTOM, , 244 p. (lire en ligne).
- (en) Christopher Mah, « Echinoderms. So What Good Are They? », sur Echinoblog, (consulté le )
- (en) Jäntschi L, Bolboaca SD, « A structural modelling study on marine sediments toxicity », Mar Drugs, vol. 6, no 2, , p. 372–88 (PMID 18728732, PMCID 2525494, DOI 10.3390/md20080017)
- (en) A Rudolph, P Medina, C Urrutia, R Ahumada, « Ecotoxicological sediment evaluations in marine aquaculture areas of Chile », Environ Monit Assess, vol. 155, nos 1-4, , p. 419–29 (PMID 18633720, DOI 10.1007/s10661-008-0444-x)
- (en) O Rulon, « The modification of developmental patterns in Arbacia eggs with malonic acid », Anat. Rec., vol. 99, no 4, , p. 652 (PMID 18895450)
- (en) J Kanungo, « Prolonged incubation in seawater induces a DNA-dependent protein phosphorylation activity in Arbacia punctulata eggs », Biochem. Biophys. Res. Commun., vol. 294, no 3, , p. 667–71 (PMID 12056821, DOI 10.1016/S0006-291X(02)00539-9, lire en ligne)
- (en) PM Failla, « RECOVERY FROM DIVISION DELAY IN IRRADIATED GAMETES OF ARBACIA PUNCTULATA », Radiat. Res., vol. 25, no 2, , p. 331–40 (PMID 14295124, DOI 10.2307/3571975, lire en ligne)
- (en) MI Sachs, E Anderson, « A cytological study of artificial parthenogenesis in the sea urchin Arbacia punctulata », J. Cell Biol., vol. 47, no 1, , p. 140–58 (PMID 4327513, PMCID 2108410, DOI 10.1083/jcb.47.1.140, lire en ligne)
- (en) GL Kite, « THE NATURE OF THE FERTILIZATION MEMBRANE OF THE EGG OF THE SEA URCHIN (ARBACIA PUNCTULATA) », Science, vol. 36, no 930, , p. 562–564 (PMID 17812420, DOI 10.1126/science.36.930.562-a, lire en ligne)
- (en) AM Zimmerman, D Marsland, « CELL DIVISION: EFFECTS OF PRESSURE ON THE MITOTIC MECHANISMS OF MARINE EGGS (ARBACIA PUNCTULATA) », Exp. Cell Res., vol. 35, , p. 293–302 (PMID 14195437, DOI 10.1016/0014-4827(64)90096-5)
- (en) A Scott, « A cytological analysis of the effects of cyanide and 4,6-dinitro-orthocresol on the mitotic phases in Arbacia punctulata », Biol. Bull., vol. 99, no 2, , p. 362–3 (PMID 14791535)
- (en) Erica Sodergren et al., « The Genome of the Sea Urchin Strongylocentrotus purpuratus », Science, vol. 314, no 5801, , p. 941-952 (lire en ligne)