Pétale
En botanique, un pétale est une pièce florale qui entoure le système reproducteur des fleurs. Constituant l'un des éléments foliacés dont l'ensemble compose la corolle d'une fleur, il correspond à une feuille modifiée.



La pigmentation des pétales fait partie, avec le parfum et le nectar, des stratégies d'attraction des pollinisateurs : les couleurs (pas forcément vives) affichées par de grandes fleurs solitaires ou isolées ainsi que par des inflorescences de petites fleurs, sont dues à l'accumulation de pigments qui fournissent un signal optique ou produisent de la chaleur, favorisant la visite des fleurs par les pollinisateurs[1].
La corolle est la partie interne du périanthe qui comprend les deux verticilles stériles d'une fleur. Les pétales sont généralement différents des sépales. Lorsqu'ils sont d'aspect identique, on parle de tépales. Dans une fleur complète, les pétales constituent souvent la partie la plus visible et colorée qui entoure les parties fertiles. Le nombre des pétales est un indicateur de la classification des plantes : on en compte généralement quatre ou cinq (le plus souvent) chez les dicotylédones, et trois (ou un multiple de trois) chez les monocotylédones.
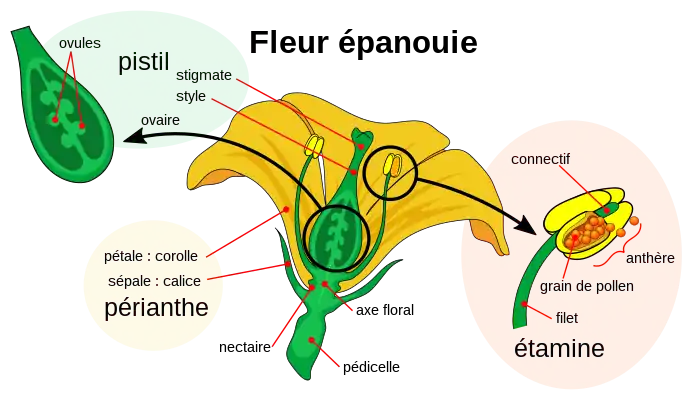
Structure
Parties d'un pétale
La diagnose distingue trois parties dans un pétale libre complet : le limbe (partie supérieure), la lame (partie intermédiaire) et l'onglet (base du pétale). Le pétale soudé dans une corolle en tube, possède une partie élargie, le limbe, et une gorge[2].
Chez les plantes de la famille des astéracées (Composées), les fleurons périphériques du capitule ont souvent un ligule très développé simulant un pétale, comme chez la marguerite par exemple. En réalité ces fleurons, qui sont les fleurs élémentaires, ont une corolle soudée en tube à sa partie inférieure, le ligule étant une excroissance latérale.
Variations
On constate d'importantes variations de la forme des pétales parmi les plantes à fleurs. Lorsque les pétales sont soudés, la corolle est dite gamopétale. Les pétales peuvent être soudés à la base, formant un « tube floral ». Chez certaines espèces, les pétales sont presque entièrement soudés, formant une sorte d'urne ou de cloche entourant le gynécée et portant les étamines. C'est le cas par exemple des gentianes.
Les fleurs de certaines espèces soit n'ont pas de pétales du tout, soit portent des pétales très réduits. Elles sont appelées « apétales ». Des exemples de fleurs à périanthe très réduit peuvent être trouvés notamment chez les arbres à chatons (fagacées par exemple) ou chez les poacées (graminées).
Lorsque les pétales sont séparés, la corolle est dite dialypétale.
Les pétales sont généralement la partie la plus apparente de la fleur, et l'ensemble des pétales, ou corolle peut présenter une symétrie radiale ou bilatérale. Si tous les pétales sont à peu près identiques en taille et en forme, la fleur est dite régulière ou actinomorphe (ce qui signifie de forme rayonnante). C'est le cas du bouton d'or par exemple. Beaucoup de fleurs qui présentent une symétrie par rapport à un plan sont dites irrégulières ou zygomorphes. Chez les fleurs irrégulières, d'autre pièces florales peuvent également perdre leur forme régulière, mais ce sont les pétales qui montrent les plus grandes modifications par rapport à la symétrie radiale. Des exemples de fleurs zygomorphes peuvent être trouvés parmi les orchidées et les fabacées (Papilionacées).
Fonctions


Le périanthe réunit des pièces stériles à la fonction protectrice (sépales et parfois pétales) et attractives (pétales), ces derniers remplissant un rôle vexillaire (expression du botaniste italien Federico Delpino, le vexillum étant l'étendard des armées romaines)[4] pour les animaux pollinisateurs avec lesquels une adaptation réciproque plus ou moins étroite a été progressivement établie (co-évolution des fleurs et des insectes). Les pétales donnent donc le caractère phanérogame (du grec phaneros, « apparent » et gamos, « union »)[5].
Plusieurs traits des pétales ont été sélectionnés au cours de l'évolution chez les plantes à fleurs phylogénétiquement éloignées, résultant d'adaptations aux divers vecteurs de pollinisation biotiques (pollinisateurs tels que les oiseaux, les abeilles et autres insectes). La convergence de ces traits a abouti à la notion de syndrome de pollinisation. Les principaux traits sont : la coloration, la taille, la forme, le guide de nectar, les nectaires floraux (situés le plus souvent à la base des pétales), les odeurs florales (en)[6] (signal olfactif d'autant plus nécessaire que la fleur est peu visible, nocturne, distante ou cachée) qui contribuent à l'attraction d'éventuels pollinisateurs et leur guidage vers les organes reproducteurs[7] - [8].
Les fleurs entomophiles sont vivement colorées dans un spectre très large de fréquences, depuis le proche ultraviolet (350 nm) jusqu'au proche infrarouge (700 nm), alliant couleurs physiques (certains blancs, bleus ou rouges)[9] - [10] et pigmentaires (pigments chromoplastiques et vacuolaires tels que les caroténoïdes ou les anthocyanes dont la couleur varie en fonction du pH). « Les insectes voient les couleurs et montrent des préférences innées vis-à-vis d'elles, selon leur espèce, et parfois à l'intérieur de l'espèce, selon la population, la colonie d'origine[11] ».
Les interactions fleurs-insectes sont souvent présentées comme un exemple modèle de relation mutualiste mais sont rarement bien analysées d'un point de vue évolutif. Les traits floraux des pétales peuvent en effet attirer les pollinisateurs mutualistes mais aussi les animaux antagonistes, notamment les herbivores. Selon le principe de l'allocation des ressources, il existerait un compromis évolutif entre ces différents traits d'attractivité[12]. Le degré de défense à l'herbivorie pourrait résulter d'un compromis entre le bénéfice de la défense, la baisse de l'herbivorie ou sa compensation, et son coût (modification de l'intensité de la couleur, de la qualité des récompenses en nectar, de la taille des pétales, de la biosynthèse des parfums floraux)[8]. Ainsi, les plantes investissent souvent dans les pétales peu de toxines ou de tannins (eux-mêmes à la toxicité moindre[13] en raison du coût trop élevé pour ces organes caducs et à vie courte), d'où le fait que de nombreuses fleurs sont comestibles et peu astringentes[14].
Évolution et phylogénie

Les pétales ou les pièces pétaloïdes ont une diversité morphologique et un développement tels qu'ils ont conduit à l'hypothèse qu'ils ne sont pas homologues mais ont évolué plusieurs fois. Cette homoplasie se retrouve dans la génétique qui montre, conformément au modèle ABC de développement floral (en) et au MADS-box, que les sépales, pétales, étamines et carpelles sont des versions modifiées les unes des autres. Il semble que les mécanismes de formation des pétales aient évolué très peu de fois (certains modèles proposent une seule origine évolutive, d'autres 6 origines différentes à partir d'un périanthe indifférencié comme celui de Amborella trichopoda[15]), plutôt que d'avoir évolué indépendamment des étamines comme chez un grand nombre de plantes[16]. La « bractée pétaloïde » de Poinsettia est analogue au pétale qui, lui, a disparu au cours de l'évolution. Les staminodes des Costaceae ont fusionné et forment un labellum pétaloïde, agissant comme un pétale. La phylogénie montre que les verticilles de pièces florales ont évolué vers la pétaloïdie à partir de bractées (bractéopétaloïdie des Astéridées, Santalales) ou d'étamines (andropétaloïdie des Caryophyllales, Rosales)[17]. La plupart des protoangiospermes possèdent un périanthe indifférencié dont les tépales montrent des gradations depuis l'extérieur avec des caractères sépaloïdes vers l'intérieur aux caractères plus pétaloïdes[18].
Notes et références
- (en) Renee Miller, Simon J. Owens, Bjørn Rørslett, « Plants and colour: Flowers and pollination », Optics & Laser Technology, vol. 43, no 2, , p. 286.
- Pierre Crété, Précis de botanique, Masson & Cie, , p. 158.
- Vincent Albouy, Pollinisation. Le génie de la nature, Quae, , p. 33.
- Félix Auguste Joseph Plateau, Comment les fleurs attirent les insectes, F. Hayez, , p. 3.
- Jean-Claude Roland, Françoise Roland, François Bouteau, Hayat El Maarouf Bouteau, Atlas de biologie végétale, Dunod, (lire en ligne), p. 129.
- La corolle, formée par l'ensemble des pétales, soudés ou non, est le principal attracteur visuel et olfactif des fleurs. . Des centaines de composés organiques volatils « qui intéressent la parfumerie ont été identifiés dans trois groupes principaux, les phénylpropanoïdes, les dérivés d'acides gras et les terpénoïdes ». Nicolas Sauvion, Paul-André Calatayud, Denis Thiéry et Frédéric Marion-Poll, Interactions insectes-plantes, éditions Quæ, (lire en ligne), p. 306
- (en) P.G. Kevan & H.G. Baker, « Insects as Flower Visitors and Pollinators », Annual Review of Entomology, vol. 28, no 1, , p. 407-453 (DOI 10.1146/annurev.en.28.010183.002203).
- (en) Danny Kessler et al., « How scent and nectar influence floral antagonists and mutualists », eLife, (DOI 10.7554/eLife.07641).
- (en) Yoseph Bar-Cohen, Biomimetics. Nature-Based Innovation, CRC Press, (lire en ligne), p. 319-325.
- (en) Jair E. Garcia, Mani Shrestha, Scarlett Howard, Phred Petersen, « Signal or cue: the role of structural colors in flower pollination », Current Zoology, vol. 65, no 4, , p. 467–481 (DOI 10.1093/cz/zoy096).
- Nicolas Sauvion, op. cit., p. 306.
- Thierry Lefevre, Michel Raymond et Frédéric Thomas, Biologie évolutive, De Boeck Superieur, (lire en ligne), p. 568.
- Ils ont une affinité moindre pour les protéines digestives des herbivores.
- Marc-André Selosse, Les Goûts et les couleurs du monde : Une histoire naturelle des tannins, de l'écologie à la santé, Actes Sud Nature, , p. 89.
- (en) Zanis MJ, « Phyolgenetic analyses and perianth evolution in basal angiosperms », Annals of the Missouri Botanical Garden, vol 90, 2003, p. 129-150.
- (en) Rasmussen, D. A.; Kramer, E. M.; Zimmer, E. A, « One size fits all ? Molecular evidence for a commonly inherited petal identity program in Ranunculales », American Journal of Botany, vol. 96, (DOI 10.3732/ajb.0800038).
- (en) LP. Ronse De Craene, « Are petals sterile stamens or bracts ? The origin and evolution of petals in the core eudicots », Ann Bot., vol. 100, no 3, , p. 621-630
- (en) Endress PK, Doyle JA, « Reconstructing the ancestral Angiosperm flower and its initial specializations », American Journal of Botany, vol 96, 2009, p. 22-66.
- Pour conserver cette couleur, les jardiniers ajoutent de l'ardoise pilée ou du sulfate d'aluminium au pied des hortensias
- Pour conserver cette couleur, les jardiniers ajoutent de l'ardoise pilée ou du sulfate d'aluminium au pied des hortensias