Miro à front rouge
Petroica goodenovii

| Règne | Animalia |
|---|---|
| Embranchement | Chordata |
| Sous-embr. | Vertebrata |
| Classe | Aves |
| Ordre | Passeriformes |
| Famille | Petroicidae |
| Genre | Petroica |
LC [1] : Préoccupation mineure
Le Miro à front rouge (Petroica goodenovii) est une espèce d'oiseaux de la famille des Petroicidae. Cette espèce est endémique d'Australie. On la trouve dans les régions sèches de la plus grande partie du continent australien où elle habite les zones arbustives et les bois clairsemés. Comme beaucoup d'oiseaux vivement colorés de la famille des Petroicidae, il présente un dimorphisme sexuel. Il mesure de 10 à 12 cm de longueur, a un petit bec mince et noir, les yeux et les pattes bruns. Le mâle a le dessus de la tête et la poitrine rouge, le dessus du corps noir et la queue noire avec l'extrémité blanche. Le ventre et les épaules sont blancs. La femelle est gris-brun. Cette espèce utilise nombre de chants variés et les mâles chantent généralement pour annoncer l'arrivée sur leur territoire et attirer les femelles. Les oiseaux se rencontrent par couples ou en petits groupes, mais le comportement social a été peu étudié.
Sa position et celle des espèces australiennes apparentées sur l'arbre de la famille des passereaux n'est pas claire ; les Petroicidae ne sont pas étroitement apparentés aux Muscicapidae européens et américains, mais semblent émaner d'un groupe primitif de Passeri. Cet oiseau se nourrit essentiellement au sol d'insectes et d'araignées. Bien qu'ayant une vaste aire de répartition, il est rare dans la majeure partie de son territoire et a reculé dans certaines régions à l'activité humaine importante.
Description
Plus petite espèce du genre Petroica, le Miro à front rouge mesure 10 à 12 cm de long avec une envergure de 15 à 19,5 cm et pèse environ 9,7 g. Mâles et femelles sont de taille similaire. Il a les pattes plus longues que les autres Petroica. Le mâle a un capuchon et la poitrine d'un rouge écarlate. Ses parties supérieures sont noir de jais avec des barres blanches sur les épaules. Sa queue est noire avec les pointes blanches. Le ventre est blanc. Toutes les couleurs sont nettement délimitées les unes par rapport aux autres[2]. La femelle est gris-brun dessus avec la couronne teintée de rougeâtre ; elle est plus pâle en dessous. Ses ailes sont brunes avec une tache chamois. Certaines femelles ont une tache rougeâtre sur la poitrine. Les deux sexes ont un petit bec noir[3], les yeux et les pattes bruns. Les oiseaux immatures ressemblent aux femelles[4] ; c'est seulement à leur seconde mue, qui a lieu à environ un an ou un peu plus, que les mâles adoptent leur plumage adulte caractéristique[5]. La mue du plumage a lieu une fois par an, après la saison de reproduction qui se déroule entre décembre et avril[6].
 Mâle adulte montrant ses épaules blanches.
Mâle adulte montrant ses épaules blanches. Femelle (ou immature).
Femelle (ou immature). Miro à front rouge en Nouvelle-Galles du Sud. Juillet 2020.
Miro à front rouge en Nouvelle-Galles du Sud. Juillet 2020.
Deux pigments caroténoïdes rouges, la canthaxanthine et l'adonirubine, sont responsables de la couleur rouge du plumage de l'oiseau[7]. Les oiseaux sont incapables de synthétiser ces composés eux-mêmes et ont besoin de les trouver dans leur nourriture. Les caroténoïdes, difficiles à métaboliser, sont également nécessaires à leur immunité : ces oiseaux doivent donc être en bonne santé pour pouvoir les utiliser aussi dans leurs plumes rouges. Le rouge du plumage est une bonne mise en valeur pour des partenaires potentiels[8]. Une étude de 2001 au parc national Terrick Terrick au Victoria a montré que les mâles qui avaient le plus de succès en matière de reproduction et étaient en meilleure condition acquéraient un plumage plus voyant lors de la mue de l'année suivante. Toutefois, l'âge des mâles et leur état apparent à l'époque de la reproduction sont plus susceptibles de prédire le succès d'accouplement pour la saison de reproduction suivante[7]. Les mâles adultes peuvent se reproduire à partir de l'âge d'un an alors qu'ils n'ont pas encore leur plumage nuptial[9] mais ils ont moins de succès pour se reproduire à cet âge[7]. Le plus âgé des oiseaux retrouvé était un oiseau bagué âgé de 5 ans et 7 mois trouvé près de Beverley, en Australie-Occidentale, en 1990[10].
On a enregistré différents cris chez eux, décrits comme des tintements et des piaillements. Ces vocalisations sont identiques sur tout le continent australien, mais diffèrent sur l'île Rottnest, sur laquelle les oiseaux enchaînent rarement leurs chants[11].
Cette espèce peut être confondue avec le Miro embrasé (P. phoenicea) et le Miro boodang (P. boodang), mais le mâle se distingue par sa couronne rouge (alors qu'elle est blanche chez les deux autres espèces) et sa petite taille ; en outre, les parties supérieures du Miro à front rouge mâle sont plus gris foncé que noires[12]. Les femelles et les oiseaux immatures sont plus difficiles à distinguer, mais ils peuvent être différenciés par la couleur rougeâtre de leur couronne et leur dessous blanc[12].
Écologie et comportement
Alimentation
.jpg.webp)
Son régime alimentaire se compose d'insectes et d'autres petits arthropodes[4]. Une étude de ses fèces effectuée près de Kambalda, en Australie-Occidentale, a révélé que 96 % de son régime alimentaire était constitué de coléoptères, les fourmis constituant la majorité du reste[13]. Parmi les autres proies retrouvées figurent des araignées et des insectes comme des sauterelles, dont le criquet australien (Chortoicetes terminifera), des chenilles et des imagos de papillons diurnes et nocturnes, notamment des Geometridae, des libellules et des demoiselles, des mantes, des fourmis-lions, des punaises, y compris des punaises de la famille des Lygaeidae et des Pentatomoidae, des perce-oreilles et des mouches (Calliphoridae, Tabanidae)[9] - [14].
Il attrape la plupart de ses proies sur le sol, mais il peut les capturer en vol. Plus rarement, il se nourrit en cherchant ses proies dans les branches basses des arbustes presque toujours à moins de 3 m au-dessus du sol. Il peut utiliser une branche basse comme point d'observation[15].
Comportement
Le Miro à front rouge vit généralement seul ou en couple[9], mais on peut voir des groupes comptant jusqu'à huit oiseaux — un couple et ses petits — en automne et en hiver[9]. Il peut former des groupes mixtes avec d'autres petits passereaux insectivores, comme la Rhipidure hochequeue (Rhipidura leucophrys), la Gérygone blanchâtre (Aphelocephala leucopsis), le Siffleur itchong (Pachycephala rufiventris) et le Langrayen gris (Artamus cinereus) au Queensland[16] et l'Acanthize à croupion roux (Acanthiza uropygialis), l'Acanthize à croupion beige (A. reguloides) ou l'Acanthize troglodyte (A. apicalis) en Australie-Occidentale[17].
Le Miro à front rouge se perche le plus souvent dans un endroit bien en vue au ras du sol, agitant souvent les ailes et la queue. Il est très actif et ne reste pas longtemps en place[18]. La femelle n'est pas très farouche mais le mâle est plus prudent avec les humains[19].
Le Miro à front rouge est territorial pendant la saison de reproduction, son territoire s'étendant de 0,25 à 1,2 ha[9]. Un couple vit et prospecte sur son territoire avant de se séparer à l'automne[9]. Le mâle défend cette zone en chantant haut et fort sur un perchoir situé à la limite de son territoire, et affronte les mâles qui pourraient y faire une incursion par un grondement dur[20]. On a vu deux mâles face à face, entre 30 cm à 1 m de distance, agitant les ailes en prenant une posture agressive tandis que la femelle couvait ses œufs[18]. On a constaté que les deux sexes réagissent à l'écoute de chants enregistrés sur bande sonore[21]. Les mâles se défendent aussi contre les incursions des Miros boodangs mâles, et inversement ils évitent d'aller chercher leur nourriture là où vit cette dernière espèce[20]. La plupart des jeunes ne peuvent pas vivre sur les territoires occupés par des oiseaux adultes et doivent trouver des terrains inoccupés. La dispersion la plus importante enregistrée à ce jour a été de 36 km, du parc national Terrick Terrick jusqu'à la forêt de Gunbower dans le nord de l'État de Victoria[22].
Reproduction
La saison de reproduction dure plus de cinq mois entre août et janvier et le couple élève jusqu'à trois couvées. Le mâle propose des sites de nidification possibles à la femelle en frottant son corps sur une fourche d'arbre qui lui semble appropriée tout en faisant des trilles. Il peut proposer plusieurs sites avant que la femelle ne décide du lieu où le nid sera établi, nid qu'elle construit seule[23]. Le nid est une coupe profonde faite d'herbes sèches et d'écorces. Des toiles d'araignées, des plumes et des poils sont utilisés pour assurer les liaisons ou le rembourrage et le nid est généralement placé à la fourche d'un arbre ou même dans un bouquet de gui[24]. Il peut être décoré avec des lichens pour se fondre dans l'environnement[25]. La femelle pond sur quelques jours consécutifs deux à trois œufs blancs teintés de bleu, de gris ou de marron et tachetés de gris-brun mesurant 16 mm × 13 mm[23] - [26]. Seule la femelle développe une plaque incubatrice et couve, mais les deux parents nourrissent les jeunes[23]. Le mâle surveille le nid perché sur une branche voisine, plutôt qu'assis sur les œufs lorsque la femelle est en quête de nourriture. Lorsqu'ils nourrissent les jeunes, les deux parents ne restent pas longtemps sur place s'il y a des prédateurs dans le voisinage[27]. Les relations extra-conjugales sont assez fréquentes, 23 % des oisillons et 37 % des couvées ont un père biologique différent de celui qui les élève, et il existe des preuves que ce type d'accouplement est davantage susceptible de donner des mâles[28].
Comme chez tous les passereaux, les poussins sont nidicoles. Ils naissent aveugles et couverts seulement d'une mince couche de duvet. Après sept jours, ils sont capables d'étirer leurs ailes et de lisser leurs plumes, et à deux semaines, de voler[23]. Pendant au moins trois semaines après que les jeunes ont quitté le nid[23], les parents les nourrissent d'araignées et d'insectes comme des mouches et des papillons de nuit[9]. Les mâles prennent en charge l'alimentation des jeunes lorsque les femelles commencent à refaire un nid pour la couvée suivante[23]. Dans une étude de terrain près de Cooma dans le sud de la Nouvelle-Galles du Sud, on a constaté que les jeunes se dispersaient hors du territoire natal au bout de quatre à six semaines pour les couples n'ayant eu qu'une couvée dans l'an, et au bout de moins d'une semaine pour deux couples qui avaient eu deux nichées dans la saison[20]. La longue saison de reproduction et les couvées multiples sont une adaptation à un climat tempéré et au niveau élevé de prédation. Malgré cela, en moyenne, seuls deux jeunes arrivent à maturité[23].
Le Coucou des buissons (Cacomantis variolosus), le Coucou pâle (C. pallidus), le Coucou de Horsfield (Chrysococcyx basalis) et le Coucou oreillard (C. osculans) viennent pondre leurs œufs dans les nids du Miro à front rouge et sont élevés par les parents d'adoption comme s'ils étaient leurs petits[29] - [30]. On a constaté toutefois que le Miro à front rouge était particulièrement agressif vis-à-vis du Coucou de Horsfield dans le parc national de Terrick Terrick et qu'aucun nid n'avait été retrouvé parasité[23].
Répartition et habitat
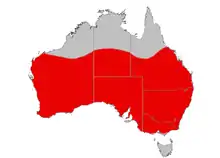
Le Miro à front rouge est présent dans toute l'Australie à l'exception de la Tasmanie, du cap York, du Top End et de la plus grande partie du Kimberley (on en trouve quelquefois dans l'extrême sud du Kimberley). Il est présent sur l'île Rottnest ainsi que sur les îles Greenly et Pearson au large de la péninsule d'Eyre[25] mais il est introuvable sur l'île Kangourou. Bien que répandu en général, il est rare dans certains endroits, comme à l'est de la Cordillère australienne, dans les régions côtières du sud du continent et dans les parties nord de son aire de répartition (il est rare d'en rencontrer au nord du 20e parallèle sud[31]). Ses déplacements sont généralement mal connus[32], en particulier en dehors de la saison de reproduction[9]. Il est sédentaire dans une grande partie du sud de son aire de répartition, bien qu'il descende au printemps et en été dans la plaine de Nullarbor, dans la région d'Adélaïde en Australie-Méridionale et au centre du Victoria. Il passe l'hiver dans le nord de son aire de répartition[29].
Le Miro à front rouge préfère les habitats plus arides que les espèces qui lui sont apparentées et habite les zones sèches tandis que le Miro boodang occupe les forêts humides où ils peuvent coexister[25]. Il habite préférentiellement les bois et les zones arbustives plantées des végétaux des genres Acacia et Callitris où dominent des espèces telles que Acacia aneura, Acacia georginae, Acacia acuminata, Callitris endlicheri, Callitris columellaris et Callitris preissii avec des arbustes en sous-étage comme les Cassia, Dodonaea, Eremophila et Triodia[32].
Taxinomie
Le Miro à front rouge a été décrit par Nicholas Aylward Vigors et Thomas Horsfield en 1827, d'après un individu collecté dans le nord du golfe Spencer dans ce qui est aujourd'hui l'Australie-Méridionale. Ils l'appellent Muscicapa goodenovii[33] et le placent parmi les Muscicapidae[34]. L'épithète spécifique goodenovii honore le révérend Samuel Goodenough, évêque de Carlisle et premier trésorier de la Société linnéenne de Londres[35].
Le Miro à front rouge est ensuite transféré dans le genre Petroica ; ce nom générique est dérivé des mots de l'ancien grec pétro, « rocher » et oikos, « maison » en raison de l'habitude qu'ont ces oiseaux de se tenir sur les rochers[36]. Dans le genre, il est l'une des cinq espèces à poitrine rouge ou rose connus en anglais sous le nom de Red Robins par opposition aux Yellow Robins du genre Eopsaltria. Ces oiseaux australiens ont été également classés pendant un certain temps dans la famille des Pachycephalidae, avant d'être placés dans leur propre famille, les Petroicidae[37]. Les études de Sibley et Ahlquist d'hybridation de l'ADN les ont placés dans l'infra-ordre des Corvida comprenant de nombreux passereaux tropicaux et australiens comme les Pardalotidae, les Maluridae et les Meliphagidae ainsi que les Corvidae[38]. Toutefois, des études de recherche moléculaire (qui font consensus) les placent comme une ramification très précoce des Passerida[39].
Il n'y aucune sous-espèce reconnue et la seule variation géographique enregistrée dans le plumage est une tendance pour les femelles vivant dans les régions les plus arides à avoir un plumage plus pâle[2]. L'étude de l'ADN nucléaire et mitochondiral des membres australiens du genre Petroica suggère que le plus proche parent du Miro à front rouge au sein de ce genre est le Miro boodang[40].
Menaces sur l'espèce
L'aire de répartition de cet oiseau étant extrêmement vaste et ses effectifs jugés suffisamment stables, l'espèce est considérée par l'Union internationale pour la conservation de la nature (UICN) comme de préoccupation mineure (LC)[41] - [1]. Cependant l'espèce a mal supporté les changements de son lieu de vie causés par l'homme. Autrefois commun dans la plaine de Cumberland dans la banlieue ouest de Sydney, elle y a maintenant presque complètement disparu[42]. Le Miro à front rouge a également disparu des environs de Rockhampton dans le Queensland et a diminué sur l'île Rottnest et dans la région de la Wheatbelt en Australie-Occidentale[32]. Des études de terrain menées dans de petits îlots de végétation restants indiquent une diminution de sa population[32].
Le chat haret est connu pour le chasser, alors que plusieurs espèces d'oiseaux dont le Corbeau d'Australie (Corvus coronoides), le Pitohui gris (Colluricincla harmonica), le Cassican à collier (Cracticus torquatus) et le Pomatostome bridé (Pomatostomus superciliosus) s'attaquent aux nids pour y prendre des jeunes[30]. Le cas d'un Méliphage à tête brune (Melithreptus brevirostris) se nourrissant d'un œuf a également été rapporté[43]. La prédation est la principale cause d'échec des couvées[23].
Annexes
Bibliographie
- (en) Walter E. Boles, The Robins and Flycatchers of Australia, Sydney, Angus & Robertson, (ISBN 0-207-15400-7)
- (en) Peter J. Higgins et Jeffrey M. Peter, Handbook of Australian, New Zealand and Antarctic Birds, vol. 6 : Pardalotes to Shrike-thrushes, Melbourne, Oxford University Press, , 1225 p. (ISBN 0-19-553762-9)
- (en) « Red-capped Robin (Petroica goodenovii) », sur Birds in Backyards, Australian Museum (consulté le )
Références taxinomiques
- (en) Référence Congrès ornithologique international : (consulté le )
- (en) Référence Zoonomen Nomenclature Resource (Alan P. Peterson) : Petroica goodenovii dans Petroicidae
- (en) Référence Tree of Life Web Project : Petroica goodenovii
- (en) Référence Catalogue of Life : Petroica goodenovii (Vigors & Horsfield, 1827) (consulté le )
- (fr+en) Référence Avibase : Petroica goodenovii (+ répartition)
- (fr+en) Référence ITIS : Petroica goodenovii (Vigors & Horsfield, 1827)
Liens externes
- (fr) Référence Oiseaux.net : Petroica goodenovii (+ répartition)
- (en) Référence Animal Diversity Web : Petroica goodenovii
- (en) Référence NCBI : Petroica goodenovii (taxons inclus)
- (en) Référence UICN : espèce Petroica goodenovii (Vigors & Horsfield, 1827) (consulté le )
Notes et références
- (en) Cet article est partiellement ou en totalité issu de l’article de Wikipédia en anglais intitulé « Red-capped Robin » (voir la liste des auteurs).
- UICN, consulté le 10 mars 2011
- Higgins et al. (2002), p. 649
- (en) K. Simpson, N. Day et P. Trusler, Field Guide to the Birds of Australia, Ringwood, Victoria, Viking O'Neil, (ISBN 0-670-90478-3), p. 174
- Boles (1988), p. 93
- Higgins et al. (2002), p. 661
- Higgins et al. (2002), p. 662
- (en) Damien K. Dowling et Raoul A. Mulder, « Red plumage and its association with reproductive success in red-capped robins », Annales Zoologici Fennici, vol. 43, no 4, , p. 311–21
- (en) V.A. Olson et I.P.F. Owens, « Costly sexual signals: are carotenoids rare, risky or required? », Trends in Ecology and Evolution, vol. 13, no 12, , p. 510–14 (DOI 10.1016/S0169-5347(98)01484-0)
- Higgins et al. (2002), p. 655
- (en) Australian Bird and Bat Banding Scheme (ABBBS), « Recovery round-up », Corella, vol. 22, , p. 126–28
- (en) M.C. Baker, E.M. Baker et M.S.A. Baker, « Songs of the Red-capped Robin, Petroica goodenovii: comparison of acoustic features in island and mainland populations », Emu, CSIRO Publishing, vol. 103, no 4, , p. 329–35 (DOI 10.1071/MU03004, lire en ligne)
- Higgins et al. (2002), p. 650
- (en) W.J. Morris et R.D. Wooller, « The structure and dynamics of an assemblage of small birds in a semi-arid eucalypt woodland in south-western Australia », Emu, vol. 101, no 1, , p. 7–12 (DOI 10.1071/MU00062, lire en ligne)
- (en) R.D. Barker et W.J.M. Vestjens, The Food of Australian Birds : (II) Passerines, Melbourne University Press, (ISBN 0-643-05115-5), p. 56–57
- Higgins et al. (2002), p. 654
- (en) Peter L. Britton, « Winter Mixed Flocks at Charters Towers, North Queensland », The Sunbird, vol. 27, no 2, , p. 29–37
- (en) E.H. Sedgwick, « Mixed associations of small birds in the south-west of Western Australia », Emu, vol. 49, no 1, , p. 9–13 (DOI 10.1071/MU949009)
- Higgins et al. (2002), p. 656
- Boles (1988), p. 90
- (en) P.P. Coventry, « Notes on the breeding behaviour of the Red-capped Robin Petroica goodenovii », Australian Bird Watcher, vol. 12, , p. 209–16
- (en) R.E. Major, F.J. Christie, G. Gowing et T.J. Ivison, « Age structure and density of Red-capped Robin populations vary with habitat size and shape », Journal of Applied Ecology, vol. 36, , p. 901–08 (DOI 10.1046/j.1365-2664.1999.00457.x, lire en ligne)
- [PDF] (en) D.K. Dowling, M. Antos et T. Sahlman, « Dispersal and recruitment of juvenile Red-capped Robins, Petroica goodenovii », Emu, vol. 203, , p. 199–205 (lire en ligne)
- (en) D.K. Dowling, « Breeding biology of the red-capped robin », Australian Journal of Zoology, CSIRO Publishing, vol. 51, no 6, , p. 533–49 (DOI 10.1071/ZO03028, lire en ligne)
- (en) S.J.N. Cooney, D.M. Watson et J. Young, « Mistletoe nesting in Australian birds: a review », Emu, CSIRO Publishing, vol. 106, , p. 1–12 (DOI 10.1071/MU04018)
- Boles (1988), p. 85
- (en) G. Beruldsen, Australian Birds : Their Nests and Eggs, Kenmore Hills, Qld, (ISBN 0-646-42798-9), p. 341–42
- Higgins et al. (2002), p. 657
- [PDF] (en) D.K. Dowling et R.A. Mulder, « Combined influence of paternal and maternal quality on sex allocation in Red-capped Robins », Journal of Evolutionary Biology, vol. 19, no 2, , p. 440–49 (PMID 16599920, DOI 10.1111/j.1420-9101.2005.01017.x, lire en ligne)
- Boles (1988), p. 86
- Higgins et al. (2002), p. 660
- Higgins et al. (2002), p. 652
- Higgins et al. (2002), p. 651
- (en) N.A. Vigors et T. Horsfield, « A description of the Australian birds in the collection of the Linnean Society; with an attempt at arranging them according to their natural affinities », Transactions of the Linnaean Society of London, vol. 15, , p. 170–331 (lire en ligne) :
« Musc. nigra ; abdomine, strigâ longitudinali alarum, rectricumque duarum lateralium marginibus albis ; fronte, pectoreque vividè coccineis. (p. 245) »
- Boles (1988), p. xv
- (en) P. Wade, Every Australian Bird Illustrated, Rigby, (ISBN 0-7270-0009-8), p. 217
- Boles (1988), p. 66
- Boles (1988), p. 35
- (en) C.G. Sibley et J.E. Ahlquist, Phylogeny and Classification of Birds : A Study in Molecular Evolution, New Haven, Yale University Press, , 603, 610–27 (ISBN 0-300-04085-7)
- [PDF] (en) F. Keith Barker, Alice Cibois, Peter A. Schikler, Julie Feinstein et Joel Cracraft, « Phylogeny and diversification of the largest avian radiation », PNAS, vol. 101, no 30, , p. 11040–45 (PMID 15263073, PMCID 503738, DOI 10.1073/pnas.0401892101, lire en ligne)
- (en) Kate Loynes, Leo Joseph et J. Scott Keogh, « Multi-locus phylogeny clarifies the systematics of the Australo-Papuan robins (Family Petroicidae, Passeriformes) », Molecular Phylogenetics and Evolution, vol. 53, no 1, , p. 212-19 (DOI 10.1016/j.ympev.2009.05.012)
- (en) « Red-capped Robin (Petroica goodenovii) », sur birdlife.org, BirdLife International (consulté le )
- (en) Peter Roberts, Birdwatcher's Guide to the Sydney Region, Kenthurst, Kangaroo Press, (ISBN 0-86417-565-5), p. 127
- (en) J.N. Hobbs, « Nest predation by two species of honeyeater », Australian Birds, vol. 24, , p. 3–4