Guyanancistrus nassauensis
Guyanancistrus nassauensis est une espèce de poissons-chats de la famille des Loricariidae. Ce petit poisson brun, indistinctement tacheté de brun-orange, est le plus petit représentant de son genre, Guyanancistrus, les plus grands individus connus mesurant environ 6 centimètres. Il se distingue des espèces proches notamment par sa grande bouche, ainsi que par ses couleurs et d'autres caractères morphologiques. Son écologie n'est pas connue, mais il semble adapté aux petits cours d'eau, aux eaux fraîches et à faible productivité. G. nassauensis est une espèce rare, hyper-endémique ne se trouvant que dans quelques cours d'eau des monts Nassau au Suriname.
.png.webp)
Ce poisson-chat est découvert en 2005 lors d'une évaluation biologique des plateaux des monts Lely et Nassau, qui montre une grande biodiversité et un fort endémisme chez les poissons de la zone. L'espèce est décrite de manière formelle en 2018, par un ichtyologiste du Suriname qui a découvert l'espèce, et deux autres du Muséum d'histoire naturelle de Genève où est notamment conservé l'holotype. Les reconstructions phylogénétiques la rapprochent de l'espèce Guyanancistrus brevispinis. Elle est menacée d'extinction par les activités minières, l'orpaillage illégal et un projet de mine de bauxite prévu dans les monts Nassau.
Description
Morphologie
.png.webp)
Guyanancistrus nassauensis est une petite espèce, le plus grand spécimen observé ayant une longueur standard de 61,0 mm. La tête et le corps sont larges et déprimés dorso-ventralement. Le profil dorsal est convexe de la pointe du museau jusqu'à l'origine de la nageoire dorsale, généralement plus aplati en arrière de l'orbite, légèrement convexe et incliné entre l'origine de la nageoire dorsale et le début de la nageoire adipeuse, légèrement concave jusqu'aux rayons de la nageoire caudale, puis remonte le long de la nageoire caudale. Le profil ventral est plat du museau jusqu'à la base de la nageoire caudale[1].
Sur la tête, il y a une carène médiane peu élevée de la pointe du museau jusqu'aux narines, parfois bordée par une dépression latérale, et une légère élévation antérieure aux orbites, l'os supra-occipital est légèrement convexe ou plat. La marge dorsale est doucement aplatie à partir de la base du premier rayon ramifié de la dorsale jusqu'à la base de la nageoire adipeuse, entre de très légères crêtes formées de chaque côté par les plaques de la série dorsale. Les premières plaques latérales de la série mi-ventrale forment une crête latérale peu élevée. Le pédoncule caudal est à peu près ovoïde en section transversale, aplati ventralement, et plus comprimé dans sa partie postérieure[1].
.png.webp)
Le museau est grand, arrondi et aplati latéralement. Les yeux sont relativement petits. La bouche est grande, ovale, avec une lèvre inférieure large, n'atteignant pas ou tout juste la ceinture pectorale, et une lèvre supérieure plus étroite. Les lèvres forment un disque ovale, couvert de courtes papilles. Une seule papille buccale, étroite, est présente, et les barbillons maxillaires sont très courts. Les dents sont courtes et fortes, avec une couronne bicuspide relativement longue, et un lobe latéral faisant environ la moitié de la taille du lobe médian[2].
La partie dorsale de la tête et du corps est munie de plaques, généralement couvertes d'odontodes courts et uniformément répartis, et la pointe du museau est en grande partie nue. La marge latérale du museau est couverte de plaques formant une armure rigide portant de courts odontodes. L'opercule porte aussi des odontodes, et une étroite zone nue borde la marge postéro-dorsale de l'opercule. Les plaques éversibles des joues portent de plus gros odontodes en nombre très variable, allant de moins de dix jusqu'à environ 35 chez certains grands spécimens. Ces odontodes jugaux sont droits avec le bout courbé, et les plus longs atteignent généralement le milieu de l'opercule, ou au-delà chez les grands spécimens. On compte deux à quatre rangées de plaques entre la plaque supra-occipitale et le spinelet de la dorsale, et la plaque nucale est souvent recouverte par la peau. Cinq séries de plaques latérales s'étendent jusqu'à la nageoire caudale. Les odontodes des séries de plaques latérales ne forment pas de carène. Les odontodes de la partie postérieure du rayon épineux de la nageoire pectorale sont un peu plus grands qu'ailleurs chez les petits spécimens, et nettement plus grands chez les spécimens de grande taille, vraisemblablement des mâles. La région abdominale est totalement nue. Il n'y a pas de structure en plaques avant la nageoire anale. La partie ventrale de la nageoire caudale est couverte de plaques montrant un nombre très réduit d'odontodes[3].
L'origine de la nageoire dorsale est légèrement antérieure à l'origine de la nageoire pelvienne. La nageoire dorsale est courte ; lorsqu'elle est appliquée, elle est loin d'atteindre la plaque impaire pré-adipeuse. La nageoire adipeuse est à peu près triangulaire, précédée par une plaque médiane (impaire), ou deux plaques fusionnées en une seule. L'épine de l'adipeuse est droite ou légèrement convexe dorsalement, et la membrane se trouvant postérieurement est convexe. Les épines des pectorales sont courtes, la pointe atteignant généralement le premier quart de l'épine des pelviennes, exceptionnellement jusqu'au premier tiers chez les individus de grande taille (vraisemblablement des mâles). La nageoire anale est courte avec une épine faible et une marge convexe. La nageoire caudale est légèrement concave, avec le lobe ventral plus long que le lobe dorsal. Les formules des nageoires sont : dorsale II,7 ; pectorale I,6 ; pelvienne i,5 ; anale i,4 (rarement i,5) ; caudale i,14,I, avec les rayons épineux numérotés en chiffres romains en lettres capitales, les rayons mous simples par des chiffres romains en lettres minuscules et ceux ramifiés par des chiffres arabes[4].
Coloration
La coloration de l'espèce vivante n'est connue que d'après la photographie d'un seul individu. La couleur dorsale du corps est brune, avec des points brun-orange un peu plus clairs et mal définis. Les nageoires sont brun-orange, et les membranes des nageoires à peine pigmentées. Dans l'alcool, la partie dorsale du corps est uniformément gris-brun, la partie ventrale jaunâtre, avec généralement des plaques de mélanophores sur les parties latérales et dans la région anale, et le ventre est blanchâtre. Les rayons des nageoires sont brunâtres, avec des taches moyennes chez certains spécimens, ces taches formant ou non des bandes. La marge de la nageoire caudale est souvent brun-orange ou brun-rouge, et les membranes des nageoires ne sont généralement pas pigmentées, ou le pigment est limité aux zones limitrophes des rayons[5].
Diagnose différentielle
.png.webp)
Guyanancistrus nassauensis est morphologiquement distinct de toutes les autres espèces du genre Guyanancistrus par sa petite taille adulte (le plus grand spécimen observé avait une longueur standard de 61 mm ; la taille adulte est probablement atteinte autour des 40 mm), par un nombre réduit de rayons de la nageoire anale (4 rayons ramifiés pour G. nassauensis, en dehors de spécimens exceptionnels, contre 5 pour les autres), et par sa grosse bouche ovale comportant deux grandes zones de dents sur le dentaire et le prémaxillaire (respectivement, en pourcentage de la longueur de la tête : 24,2–31,9, en moyenne 27,6 chez G. nassauensis, contre 23,6 ou moins chez les autres espèces, sauf chez G. niger, et de 25,4–31,4, en moyenne 28,1, contre 24,5 ou moins). Seul G. niger a les dentaires presque aussi grands (22,5–26,3, en moyenne 25,0 % de la longueur de la tête), mais ses prémaxillaires sont plus courts (de 21,7–23,6, en moyenne de 22,6 % de la longueur de la tête)[1].

G. nassauensis se distingue de G. longispinis et de G. niger par un rayon épineux de la nageoire pectorale beaucoup plus court (en pourcentage de la longueur standard : 22,2–26,3, en moyenne 24,4 chez G. nassauensis, contre 31,9–45,5, en moyenne 40,2 chez G. longispinis et 33,3–48,0, en moyenne 42,8 chez G. niger), par des motifs de couleur (corps et nageoires uniformément bruns ou avec des taches de taille moyenne plus pâles et indistinctes, contre du brun-noir avec des petites taches jaunes arrondies pour G. longispinis, ou des points blancs pour G. niger). Il est différencié de toutes les espèces du groupe de G. brevispinis en ayant, en moyenne, le corps plus large, la tête la plus profonde et la plus longue, la plus grande distance interbranchiale, les plus courtes nageoires et le plus grand nombre de dents[1].
G. nassauensis se distingue de toutes les autres espèces du genre à l'exception de G. brevispinis par ses séquences spécifiques du marqueur « code-barres » qu'est le gène COI[1].
Écologie
Mis à part son habitat, peu d'informations sont disponibles sur l'écologie de Guyanancistrus nassauensis. Quelques traits de son comportement ont été décrits à partir d'observations en aquarium de quatre spécimens conservés avec des individus de l'espèce Harttiella crassicauda en . G. nassauensis était actif pendant la journée et la nuit, et ne se cachait pas à l'abri, mais restait à découvert même dans le cas d'un individu transféré dans un plus grand aquarium avec de nombreuses autres espèces de poissons. L'espèce n'était pas territoriale mais tolérait la présence d'autres espèces de poissons, conspécifiques ou pas. G. nassauensis passait essentiellement son temps sur les vitres de l'aquarium et sur les macrophytes, les débris de bois et les rochers, mais plutôt pas sur le substrat sablonneux du fond. En aquarium, G. nassauensis pouvait être nourri aux flocons végétariens après une courte période d'acclimatation, et a également été observé se nourrissant d'algues[6].
Distribution et habitat
Emplacement des monts Nassau au Suriname. |
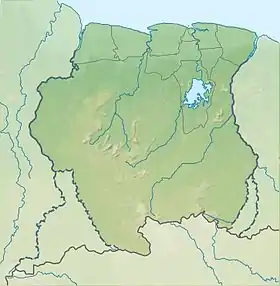
Guyanancistrus nassauensis est connu uniquement de la Paramaka Creek et certains de ses affluents, du bassin du Maroni, dans les monts Nassau au Suriname (d'une superficie d'environ 20 × 20 km2). Dans la Paramaka Creek, G. nassauensis a été trouvé en syntopie avec des juvéniles de l'espèce G. brevispinis et avec Harttiella crassicauda, une seconde espèce endémique des monts Nassau. Cependant, G. nassauensis se trouve à la fois dans les sources pérennes et dans la partie supérieure du tronçon principal de Paramaka Creek (pentes inférieures du plateau, entre 120 et 530 m d'altitude), tandis que H. crassicauda ne se trouve que sur le plateau (entre 230 et 530 m d'altitude). Dans l'IJs Creek (4° 51′ 39″ N, 54° 35′ 59″ O), affluent de la Paramaka Creek sur le plateau des Nassau (à 467 m), G. nassauensis et H. crassicauda se trouvent tous deux dans les eaux fraîches, claires et peu profondes à faible conductivité, au pH neutre, à concentrations d'azote inorganique faibles, et celles d'azote organique relativement élevées, des quantités de phosphore total faibles et de carbone organique élevées. Le substrat du fond est composé de gravier avec des rochers et de la roche (avec des algues rouges filamenteuses du genre Batrachospermum accrochées à elle), et près du bord du plateau dans des eaux un peu plus profondes (environ 50 cm) où pousse la macrophyte Thurnia sphaerocephala. Son nanisme pourrait être une adaptation aux petits ruisseaux, aux eaux fraîches et à faible productivité. Dans la partie supérieure du tronçon principal de la Paramaka Creek, ainsi que dans certaines branches en amont, sur le plateau, G. nassauensis se trouve en syntopie avec G. brevispinis[7]. La localité type est située à 277 m d'altitude dans une branche nord de la Paramaka Creek, un cours d'eau de taille moyenne peu profond (de 3 à 7 m de large et moins de 50 cm de profondeur) avec des fosses, un substrat rocheux, et bordé par le sol ferme de la forêt tropicale[5].
Taxinomie et systématique

| Relations phylogénétiques au sein du genre Guyanancistrus[8] : | |
L'espèce est découverte en 2005 par une équipe de 13 chercheurs qui explorent une région à environ 130 kilomètres au sud de Paramaribo pour réaliser l'évaluation biologique des plateaux des monts Lely et Nassau. Cette expédition montre une grande biodiversité et un fort taux d'endémisme chez les poissons de la zone. Parmi ceux-ci figurent plusieurs espèces inconnues de la science, dont une nommée « Guyanancistrus sp. "grosse bouche" » en raison de sa bouche inhabituellement grande[9] - [10] - [11]. Elle est formellement décrite en 2018 sous le nom de Guyanancistrus nassauensis, par un ichtyologiste du Suriname, Jan H. Mol, et deux chercheurs du Muséum d'histoire naturelle de Genève (MHNG), Sonia Fisch-Muller et Raphaël Covain[1]. Le genre Guyanancistrus, décrit en 2001, doit son nom à la contraction de « Guyane » et du nom de genre Ancistrus[12] ; l'épithète spécifique nassauensis est une référence à la distribution de l'espèce qui n'est connue que de cours d'eau des monts Nassau, adjoint du suffixe latin -ensis, « qui vit dans, qui habite »[5]. L'holotype de G. nassauensis, collecté en 2006, est conservé dans les collections du muséum de Genève, et des paratypes sont déposés dans l'institution genevoise, au musée d'histoire naturelle de l'université d'Auburn (AUM) et dans la Collection zoologique nationale du Suriname (NZCS)[5].
Les reconstructions phylogénétiques au sein du genre Guyanancistrus publiées en 2018 montrent qu'une lignée comprenant G. niger et G. longispinis se distingue de toutes les autres espèces. Ces dernières forment un groupe frère comprenant un clade qui regroupe G. nassauensis et G. brevispinis, quand un second rassemble les quatre autres espèces connues du genre : G. tenuis, G. megastictus, G. teretirostris et G. brownsbergensis[8].
Menaces et conservation
.png.webp)
Guyanancistrus nassauensis et G. brownsbergensis sont connus chacun dans un seul cours d'eau de montagne, respectivement dans les monts Nassau (Paramaka Creek) et dans les monts Brownsberg (Kumbu Creek). Avec cette aire de répartition très restreinte (moins de 20 × 20 km2), les deux espèces peuvent être considérées comme hyper-endémiques et sont actuellement menacées d'extinction par un projet de mine de bauxite et par les activités d'orpaillage illégal déjà en cours dans la région[7]. Comme les monts Nassau sont éloignés de toute zone habitée, ils sont totalement dénués de protection[9].
Annexes
Bibliographie
- (en) Sonia Fisch-Muller, Jan H. Mol et Raphaël Covain, « An integrative framework to reevaluate the Neotropical catfish genus Guyanancistrus (Siluriformes: Loricariidae) with particular emphasis on the Guyanancistrus brevispinis complex », PLOS One, PLoS, vol. 13, no 1, , e0189789 (ISSN 1932-6203, OCLC 228234657, PMID 29298344, PMCID 5752014, DOI 10.1371/JOURNAL.PONE.0189789, lire en ligne)
Liens externes
- Ressources relatives au vivant :
Notes et références
- Fisch-Muller, Mol & Covain (2018), p. 38.
- Fisch-Muller, Mol & Covain (2018), p. 38-39.
- Fisch-Muller, Mol & Covain (2018), p. 39.
- Fisch-Muller, Mol & Covain (2018), p. 39-40.
- Fisch-Muller, Mol & Covain (2018), p. 40.
- (en) Kenneth Wan Tong You, « Appendix 15. Observations on the behavior of Harttiella crassicauda and Guyanancistrus n.sp. (‘big mouth’) of Nassau Mountains in the aquarium », dans Leeanne E. Alonso et Jan H.A. Mol, A Rapid Biological Assessment of the Lely and Nassau Plateaus, Suriname (with additional information on the Brownsberg Plateau), vol. 43, Conservation International, coll. « RAP Bulletin of Biological Assessment », , 107-118 p. (lire en ligne).
- Fisch-Muller, Mol & Covain (2018), p. 58.
- Fisch-Muller, Mol & Covain (2018), p. 18-19.
- (en) Deborah Zabarenko, « Purple frog among 24 new species found in Suriname », (consulté le ).
- (en) Jan H.A. Mol, Kenneth Wan Tong You, I. Vrede, A. Flynn, P. Ouboter et F. Van der Lugt, « Fishes of Lely and Nassau Mountains, Suriname », dans Leeanne E. Alonso et Jan H.A. Mol, A Rapid Biological Assessment of the Lely and Nassau Plateaus, Suriname (with additional information on the Brownsberg Plateau), vol. 43, Conservation International, coll. « RAP Bulletin of Biological Assessment », , 107-118 p. (lire en ligne).
- (en) Jan H.A. Mol, The freshwater fishes of Suriname, Leiden, Netherlands, Brill, , p. 450-451.
- (de) Isaäc J.H. Isbrücker, Ingo Seidel, J. Pieter Michels, Erwin Schraml et Andrè Werner, « Diagnose vierzehn neuer Gatungen der Familie Loricariidae Rafinesque, 1815 (Teleostei, Ostariophysi) », Die Aquarien- und Terrarienzeitschrift, Stuttgart, vol. spécial no 2, , p. 17-24.