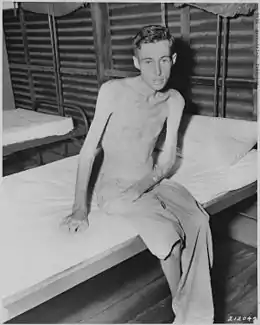
Fonte musculaire
Une fonte musculaire peut être observée dans de nombreuses situations physiologiques ou pathologiques :
- lors du vieillissement ;
- en absence d'activité physique ;
- en présence d'un hypercatabolisme ;
- lors de maladies génétiques…
| Spécialité | Rhumatologie et neurologie |
|---|
| CIM-10 | M62.5 |
|---|---|
| CIM-9 | 728.2 |
| DiseasesDB | 29472 |
| MedlinePlus | 003188 |
| MeSH | D009133 |
![]() Mise en garde médicale
Mise en garde médicale
Tous les tissus de l'organisme comme le muscle squelettique subissent un remodelage permanent permettant le remplacement des parties altérées et assurant ainsi le maintien de l'intégrité de leurs fonctions. Les protéines, dont le rôle est aussi bien contractile, structurel que métabolique, sont une composante essentielle du muscle. Leur renouvellement permanent dépend de processus biochimiques qui impliquent simultanément la synthèse et le catabolisme. L'équilibre entre synthèse et catabolisme conditionne la conservation de la masse des protéines musculaires.
Fonte musculaire liée à l'âge
Diminution de la masse musculaire liée à l'âge
Entre 20 et 60 ans on observe une diminution d'environ 40 % de la section transversale du muscle chez l'Homme[1] - [2] - [3].
La fonte musculaire avec l'avancée en âge s'explique essentiellement par une perte de fibres musculaires de type I et II[4] - [5]. Alors que le nombre de fibres baisse de seulement 5 % entre 24 et 50 ans, une diminution de 30 et 40 % est observée entre 50 et 80 ans[4]. Cela aboutit à une réduction annuelle d'environ 1 % de la surface de section du muscle au-delà de 50 ans[6] - [7].
Cependant, l'atrophie de la fibre musculaire, c'est-à-dire la diminution de leur diamètre, est également impliquée pour expliquer la diminution de la masse musculaire liée à l'âge[5] - [8] - [9]. Contrairement à la diminution de la masse musculaire liée à l'âge, l'atrophie n'affecte pas de manière similaire tous les types de fibres musculaires. En effet, ce sont les fibres de type II qui semblent être le plus affectées par l'avance en âge, avec une baisse de 20 à 60 % de leur diamètre[10] - [11].
De nombreuses hypothèses ont été avancées pour expliquer l'atrophie des fibres musculaires des populations âgées. L'une des principales causes retenues concerne la diminution de la synthèse des protéines musculaires avec l'âge. Elle touche tout particulièrement les chaînes lourdes de myosine, dont la synthèse peut diminuer de 45 % au-delà de 75 ans[12] - [13] - [14].
Diminution de la force musculaire liée à l'âge
En moyenne, la force musculaire des extenseurs du genou est réduite de 20 et 40 % entre 20 et 70-80 ans[15] - [16]. Des résultats du même ordre sont observés pour les fléchisseurs de l'épaule et du poignet[17] - [18]. La réduction de force musculaire semble s'accélérer tout particulièrement entre 60-70 ans. Des études longitudinales observent ainsi une réduction de 30 à 40 % de la force des extenseurs du genou et de l'épaule entre 60 et 70 ans[8] - [19].
Sarcopénie
Lorsque la fonte musculaire liée à l'âge atteint un certain seuil, elle conduit à une pathologie : la sarcopénie. Toutefois, il existe des stratégies nutritionnelles pouvant la prévenir.
Fonte musculaire liée à l'inactivité physique
Diminution de la masse musculaire liée à l'inactivité physique
L'immobilisation entraine une réduction de la surface et de la taille des fibres musculaire.
Chez des sujets adultes, 2 à 5 semaines d'immobilisation réduisent la surface des fibres de 10 à 70 % suivant la durée d'immobilisation et le type de fibres, les fibres de type I étant les plus touchées[20] - [21] - [22]. Après 3 semaines d'alitement ou 14 jours de suspension de la jambe, une diminution d'environ 10 % de la section transversale du quadriceps est observée[23] - [24] - [25]. Celle-ci peut atteindre 18 % après 17 semaines d'alitement[26].
La réduction de la masse musculaire due à l'inactivité s'explique principalement par une atrophie des fibres de type I et II[27] - [28] - [29]. Néanmoins, contrairement aux effets de l'avance en âge, les fibres de type I semblent davantage affectées par l'inactivité. Après 6 semaines d'alitement, plusieurs études ont montré chez l'Homme d'importantes diminutions du diamètre des fibres de type I et une diminution plus faible du diamètre des fibres de type II[22] - [30] - [31].
La réduction de l'activité physique est une situation fréquemment observée chez les personnes hospitalisées pour une longue durée mais également chez les personnes âgées, qu'elle soit liée à une faiblesse musculaire, à une chute ou à la suite d'une pathologie. L'atrophie musculaire générée par ces périodes d'immobilisation prolongées est plus importante chez un sujet âgé que chez un sujet jeune[32] - [33].
Diminution de la force musculaire liée à l'inactivité physique
Chez l'homme, une période de 1 à 2 semaines d'inactivité des membres inférieurs induit une réduction de 15 % de la force des extenseurs du genou chez des sujets d'une vingtaine d'années[34] - [35] - [36]. Cette diminution peut atteindre plus de 20 % après 4 semaines d'inactivité[37].
Fonte musculaire liée à un hypercatabolisme
L'hypercatabolisme protéique correspond à une dégradation anormalement accélérée des protéines de l'organisme. Il est l'une des caractéristiques de la réponse métabolique à l'agression (acte chirurgical, cancers, traumatisme, personnes âgées fragiles…). La balance azotée est dans cette situation toujours négative et les pertes d'azote peuvent être considérables (> 25 g·L-1, soit 500 g·j-1 de masse maigre). Les acides aminés libérés à partir des protéines musculaires sont utilisés par le foie pour la gluconéogenèse et la synthèse des protéines de la phase aiguë. Il a été récemment montré chez des traumatisés crâniens que la protéolyse totale était multipliée par 2,2 et la synthèse protéique totale par 2[38].
Fonte musculaire liée à des maladies neuronales
Les maladies neuromusculaires se traduisent pour la plupart par une perte de force musculaire transitoire (myasthénie, paralysies périodiques) ou permanente et s'aggravant progressivement (dystrophies musculaires progressives) ou restant stable (amyotrophies spinales). Cette perte de force s'accompagne le plus souvent d'une fonte musculaire (amyotrophie). Ce sont des maladies liées à une anomalie, soit de la fibre musculaire, soit de la jonction de celle-ci avec son nerf moteur (motoneurone), soit du nerf moteur (motoneurone périphérique). Il y a plus de 40 maladies liées à une atteinte primitive et isolée de l'unité motrice, que l'on peut rassembler en plusieurs groupes :
- atteintes de la fibre musculaire : myopathies ;
- atteintes de la jonction musculaire : myasthénie ;
- atteintes du nerf moteur périphérique : maladie de Charcot-Marie-Tooth.
Stratégie de prévention de la fonte musculaire
L'exercice physique
Les exercices en endurance permettant d'améliorer l'équilibre et l'appétit et d'augmenter les capacités respiratoires et la synthèse protéique musculaire en ayant toutefois une action minime sur la force musculaire[39] - [40].
La pratique d'une activité physique régulière et adaptée alliant des exercices en endurance et des exercices en force est essentielle pour ralentir le phénomène de vieillissement musculaire. Cependant, l'activité physique de loisir n'est pas suffisante pour prévenir totalement la diminution de masse musculaire avec l'âge mais elle permet de retarder la sensation de fatigue, d'améliorer l'équilibre et ainsi de prévenir les chutes et de contribuer à améliorer la qualité de vie.
Nutrition pulsée
Elle consiste à apporter 80 % des apports protéiques journaliers recommandés (soit 1 à 1,2 g·kg-1 de protéines par jour chez la personne âgée) en un seul repas, de préférence le déjeuner. Cette stratégie permet de saturer partiellement l'extraction splanchnique et ainsi d'obtenir une meilleure biodisponibilité des acides aminés pour la stimulation de la synthèse protéique postprandiale[41] - [42] - [43] - [44] - [45] - [46] - [47]. Elle est cependant difficile à mettre en œuvre.
Leucine
La leucine stimule la protéosynthèse musculaire par son action sur la voie mTOR[48] - [49] - [50]. Cependant, cet effet est limité en raison de l'augmentation de la séquestration splanchnique de la leucine avec l'âge[46]. D'autre part, il n'a pas été démontré qu'une complémentation à long terme en leucine soit efficace pour augmenter la masse musculaire chez les sujets âgés[51].
Citrulline
La citrulline est le seul acide aminé à ne pas être capté par le foie ce qui lui permet d'échapper à l'extraction splanchnique. La citrulline a un pouvoir stimulant sur la voie mTOR[52]. Ainsi, un apport chronique de L-citrulline chez le rat âgé permet de corriger les effets du vieillissement sur la composition corporelle[53] et notamment d'augmenter de l'ordre de 20 % la masse musculaire[53]. Ce travail expérimental a été confirmé chez l'Homme puisqu'il apparaît qu'un apport de citrulline par voie orale permet d'augmenter de 25 % la vitesse de synthèse protéique musculaire[54].
Vitamine D
La vitamine D a montré son efficacité dans la réduction du nombre de chutes liées à la faiblesse musculaire[55]. Il est conseillé chez les personnes âgées de consommer 800 UI de vitamine D3 par jour (soit 20 µg).
« Protéines rapides »
La vitesse d'absorption des protéines peut jouer un rôle important dans la biodisponibilité post prandiale des acides aminés et donc avoir une incidence sur la stimulation de la synthèse protéique musculaire[56]. Dans ce contexte, plus l'apparition des acides aminés dans le sang est massive et rapide, plus l'aire splanchnique a des chances d'être saturée[57]. Il reste à déterminer si l'effet se poursuit à long terme et surtout si la consommation de « protéines rapides » (comme les protéines du lactosérum) a une incidence sur la masse et la force musculaire ainsi que sur la performance physique.
Notes et références
- (en) Doherty TJ, Vandervoort AA, Brown WF. « Effects of ageing on the motor unit: a brief review » Can J Appl Physiol. 1993 déc;18(4):331-58.
- (en) Vandervoort AA. « Aging of the human neuromuscular system » Muscle Nerve 2002 janv;25(1):17-25.
- (en) Klitgaard H, Mantoni M, Schiaffino S, Ausoni S, Gorza L, Laurent-Winter C. et al. « Function, morphology and protein expression of ageing skeletal muscle: a cross-sectional study of elderly men with different training backgrounds » Acta Physiol. Scand. 1990 sept;140(1):41-54.
- (en) Aniansson A, Grimby G, Hedberg M. « Compensatory muscle fiber hypertrophy in elderly men » J. Appl. Physiol. 1992 sept;73(3):812-6.
- (en) Lexell J, Taylor CC, Sjöström M. « What is the cause of the ageing atrophy? Total number, size and proportion of different fiber types studied in whole vastus lateralis muscle from 15- to 83-year-old men » J. Neurol. Sci. 1988 avr;84(2-3):275-94.
- (en) Kent-Braun JA, Ng AV. « Specific strength and voluntary muscle activation in young and elderly women and men » J. Appl. Physiol. 1999 juill;87(1):22-9.
- (en) Frontera WR, Reid KF, Phillips EM, Krivickas LS, Hughes VA, Roubenoff R. et al. « Muscle fiber size and function in elderly humans: a longitudinal study » J. Appl. Physiol. 2008 août;105(2):637-42.
- (en) Aniansson A, Hedberg M, Henning GB, Grimby G. « Muscle morphology, enzymatic activity, and muscle strength in elderly men: a follow-up study » Muscle Nerve 1986 sept;9(7):585-91.
- (en) Lexell J, Downham D. « What determines the muscle cross-sectional area? » J. Neurol. Sci. 1992 août;111(1):113-4.
- (en) Larsson L, Sjödin B, Karlsson J. « Histochemical and biochemical changes in human skeletal muscle with age in sedentary males, age 22--65 years » Acta Physiol. Scand. 1978 mai;103(1):31-9.
- (en) Essén-Gustavsson B, Borges O. « Histochemical and metabolic characteristics of human skeletal muscle in relation to age » Acta Physiol. Scand. 1986 janv;126(1):107-14.
- (en) Balagopal P, Rooyackers OE, Adey DB, Ades PA, Nair KS. « Effects of aging on in vivo synthesis of skeletal muscle myosin heavy-chain and sarcoplasmic protein in humans » Am. J. Physiol. 1997 oct;273(4 Pt 1):E790-800.
- (en) Balagopal P, Schimke JC, Ades P, Adey D, Nair KS. « Age effect on transcript levels and synthesis rate of muscle MHC and response to resistance exercise » Am. J. Physiol. Endocrinol. Metab. 2001 févr;280(2):E203-208.
- (en) Short KR, Vittone JL, Bigelow ML, Proctor DN, Coenen-Schimke JM, Rys P. et al. « Changes in myosin heavy chain mRNA and protein expression in human skeletal muscle with age and endurance exercise training » J. Appl. Physiol. 2005 juill;99(1):95-102.
- (en) Larsson L, Grimby G, Karlsson J. « Muscle strength and speed of movement in relation to age and muscle morphology » J Appl Physiol. 1979 mars;46(3):451-6.
- (en) Murray MP, Duthie EH Jr, Gambert SR, Sepic SB, Mollinger LA. « Age-related differences in knee muscle strength in normal women » J Gerontol. 1985;40(3):275-80.
- (en) McDonagh MJ, White MJ, Davies CT. « Different effects of ageing on the mechanical properties of human arm and leg muscles » Gerontology 1984;30(1):49-54.
- (en) Bassey EJ, Harries UJ. « Normal values for handgrip strength in 920 men and women aged over 65 years, and longitudinal changes over 4 years in 620 survivors » Clin. Sci. 1993;84(3):331-7.
- (en) Frontera WR, Hughes VA, Fielding RA, Fiatarone MA, Evans WJ, Roubenoff R. « Aging of skeletal muscle: a 12-yr longitudinal study » J. Appl. Physiol. 2000;88(4):1321-6.
- (en) Thomason DB, Booth FW. « Atrophy of the soleus muscle by hindlimb unweighting » J. Appl. Physiol. 1990 janv;68(1):1-12.
- (en) Nicks DK, Beneke WM, Key RM, Timson BF. « Muscle fibre size and number following immobilisation atrophy » J. Anat. 1989 avr;163:1-5.
- (en) Berg HE, Larsson L, Tesch PA. « Lower limb skeletal muscle function after 6 wk of bed rest » J. Appl. Physiol. 1997 janv;82(1):182-8.
- (en) Funato K, Matsuo A, Yata H, Akima H, Suzuki Y, Gunji A, et al. « Changes in force-velocity and power output of upper and lower extremity musculature in young subjects following 20 days bed rest » J Gravit Physiol. 1997 janv;4(1):S22-30.
- (en) Kawakami Y, Muraoka Y, Kubo K, Suzuki Y, Fukunaga T. « Changes in muscle size and architecture following 20 days of bed rest » J Gravit Physiol. 2000 déc;7(3):53-9.
- (en) Jones SW, Hill RJ, Krasney PA, O'Conner B, Peirce N, Greenhaff PL. « Disuse atrophy and exercise rehabilitation in humans profoundly affects the expression of genes associated with the regulation of skeletal muscle mass » FASEB J. 2004 juin;18(9):1025-7.
- LeBlanc AD, Schneider VS, Evans HJ, Pientok C, Rowe R, Spector E. « Regional changes in muscle mass following 17 weeks of bed rest » J. Appl. Physiol. 1992;73(5):2172-8.
- (en) Allen DL, Yasui W, Tanaka T, Ohira Y, Nagaoka S, Sekiguchi C. et al. « Myonuclear number and myosin heavy chain expression in rat soleus single muscle fibers after spaceflight » J. Appl. Physiol. 1996;81(1):145-51.
- (en) Fitts RH, Romatowski JG, De La Cruz L, Widrick JJ, Desplanches D. « Effect of spaceflight on the maximal shortening velocity, morphology, and enzyme profile of fast- and slow-twitch skeletal muscle fibers in rhesus monkeys » J Gravit Physiol. 2000 janv;7(1):S37-38.
- (en) Baldwin KM, Haddad F. « Effects of different activity and inactivity paradigms on myosin heavy chain gene expression in striated muscle » J. Appl. Physiol. 2001 janv;90(1):345-57.
- (en) Ferretti G, Antonutto G, Denis C, Hoppeler H, Minetti AE, Narici MV. et al. « The interplay of central and peripheral factors in limiting maximal O2 consumption in man after prolonged bed rest » J. Physiol. (Lond.) 1997;501( Pt 3):677-86.
- (en) Borina E, Pellegrino MA, D'Antona G, Bottinelli R. « Myosin and actin content of human skeletal muscle fibers following 35 days bed rest » Scand J Med Sci Sports. 2010;20(1):65-73.
- (en) Kortebein P, Ferrando A, Lombeida J, Wolfe R, Evans WJ. « Effect of 10 days of bed rest on skeletal muscle in healthy older adults » JAMA 2007;297(16):1772-4.
- (en) Park H, Park S, Shephard R, Aoyagi Y. « Yearlong physical activity and sarcopenia in older adults: the Nakanojo Study » European Journal of Applied Physiology 2010;109(5):953-61.
- (en) Berg HE, Tesch PA. « Changes in muscle function in response to 10 days of lower limb unloading in humans » Acta Physiol. Scand. 1996;157(1):63-70.
- (en) Deschenes MR, Holdren AN, McCoy RW. « Adaptations to short-term muscle unloading in young and aged men » Med Sci Sports Exerc. 2008;40(5):856-63.
- (en) Suetta C, Hvid LG, Justesen L, Christensen U, Neergaard K, Simonsen L. et al. « Effects of aging on human skeletal muscle after immobilization and retraining » J. Appl. Physiol. 2009;107(4):1172-80.
- (en) Berg HE, Dudley GA, Häggmark T, Ohlsén H, Tesch PA. « Effects of lower limb unloading on skeletal muscle mass and function in humans » J. Appl. Physiol. 1991;70(4):1882-5.
- (en) Flakoll PJ, Wentzel LS, Hyman SA. « Protein and glucose metabolism during isolated closed-head injury » Am. J. Physiol. 1995;269(4 Pt 1):E636-641.
- (en) Onambélé-Pearson GL, Breen L, Stewart CE. « Influence of exercise intensity in older persons with unchanged habitual nutritional intake: skeletal muscle and endocrine adaptations » Age (Dordr) 2010;32(2):139-53.
- (en) Latham N, Anderson C, Bennett D, Stretton C. « Progressive resistance strength training for physical disability in older people » Cochrane Database Syst Rev. 2003;(2):CD002759.
- Cynober L, Alix E, Arnaud-Battandier F, Bonnefoy M, Brocker P, Cals M-J. et al. « Apports nutritionnels conseillés chez la personne âgée » Nutrition Clinique et Métabolisme 2000;14(S1):3-60.
- (en) Arnal M-A, Mosoni L, Dardevet D, Ribeyre M-C, Bayle G, Prugnaud J. et al. « Pulse protein feeding pattern restores stimulation of muscle protein synthesis during the feeding period in old rats » J. Nutr. 2002;132(5):1002-8.
- (en) Boirie Y, Gachon P, Beaufrère B. « Splanchnic and whole-body leucine kinetics in young and elderly men » Am. J. Clin. Nutr. 1997;65(2):489-95.
- (en) Jourdan M, Cynober L, Moinard C, Blanc MC, Neveux N, De Bandt JP. et al. « Splanchnic sequestration of amino acids in aged rats: in vivo and ex vivo experiments using a model of isolated perfused liver ». Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008;294(3):R748-755.
- (en) Guillet C, Prod'homme M, Balage M, Gachon P, Giraudet C, Morin L. et al. « Impaired Anabolic Response of Muscle Protein Synthesis Is Associated with S6K1 Dysregulation in Elderly Humans » FASEB J [Internet] 2004
- Cynober L, Jourdan M, Aussel C, Guillet C, Walrand S, Boirie Y. « Sarcopénie des sujets âgés : libérez les acides aminés ! » Nutrition Clinique et Métabolisme 2004;18(4):198-204.
- (en) Arnal MA, Mosoni L, Boirie Y, Houlier ML, Morin L, Verdier E. et al. « Protein pulse feeding improves protein retention in elderly women » Am. J. Clin. Nutr. 1999 juin;69(6):1202-8.
- (en) Guillet C, Zangarelli A, Mishellany A, Rousset P, Sornet C, Dardevet D. et al. « Mitochondrial and sarcoplasmic proteins, but not myosin heavy chain, are sensitive to leucine supplementation in old rat skeletal muscle » Exp. Gerontol. 2004;39(5):745-51.
- (en) Rieu I, Sornet C, Bayle G, Prugnaud J, Pouyet C, Balage M. et al. « Leucine-supplemented meal feeding for ten days beneficially affects postprandial muscle protein synthesis in old rats » J. Nutr. 2003;133(4):1198-205.
- (en) Rieu I, Balage M, Sornet C, Giraudet C, Pujos E, Grizard J. et al. « Leucine supplementation improves muscle protein synthesis in elderly men independently of hyperaminoacidaemia » J. Physiol. (Lond.) 2006;575(Pt 1):305-15.
- (en) Verhoeven S, Vanschoonbeek K, Verdijk LB, Koopman R, Wodzig WKWH, Dendale P. et al. « Long-term leucine supplementation does not increase muscle mass or strength in healthy elderly men » Am. J. Clin. Nutr. 2009;89(5):1468-75.
- (en) Le Plénier S, Walrand S, Noirt R, Cynober L, Moinard C. « Effects of leucine and citrulline versus non-essential amino acids on muscle protein synthesis in fasted rat: a common activation pathway? » Amino Acids 1-8.
- (en) Moinard C. « Long-term effect of citrulline supplementation in healthy aged rats: effect on body composition » Clinical Nutrition Supplements 2009;4(2):12.
- (en) Jourdan M, Nair KS, Ford C, Shimke J, Ali B, Will B. et al. « Citrulline stimulates muscle protein synthesis at the post-absoptive state in healthy subjects fed a low-protein diet » Clinical Nutrition Supplements 2008;3:11-2.
- (en) Bischoff-Ferrari HA, Dawson-Hughes B, Staehelin HB, Orav JE, Stuck AE, Theiler R. et al. « Fall prevention with supplemental and active forms of vitamin D: a meta-analysis of randomised controlled trials » BMJ 2009;339:b3692.
- (en) Boirie Y, Dangin M, Gachon P, Vasson MP, Maubois JL, Beaufrère B. « Slow and fast dietary proteins differently modulate postprandial protein accretion » Proc. Natl. Acad. Sci. U.S.A. 1997;94(26):14930-5.
- (en) Dangin M, Guillet C, Garcia-Rodenas C, Gachon P, Bouteloup-Demange C, Reiffers-Magnani K. et al. « The rate of protein digestion affects protein gain differently during aging in humans » J. Physiol. (Lond.) 2003;549(Pt 2):635-44.