Thalamus humain
Le thalamus (du grec θαλαμος : « chambre ») constitue un volumineux noyau ovoïde, d'origine diencéphalique, pair et symétrique de chaque côté du troisième ventricule.
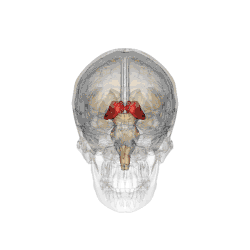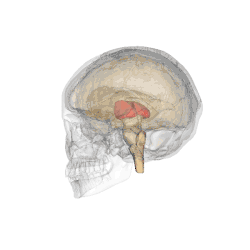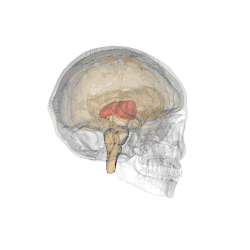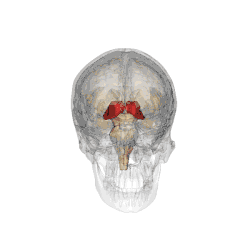
Il se subdivise en différents sous-noyaux : antérieur, ventraux, dorso-latéraux, dorso-médian, intralaminaires, postérieurs (incluant le pulvinar) et genouillés. Les deux-tiers antérieurs de sa face externe est recouverte par le noyau réticulaire.
D'un point de vue morphologique, il entre globalement en rapport avec : 1. en dehors, la capsule interne, 2. en bas, la région sous-thalamique (zona incerta), 3. en dedans, le système ventriculaire latéral et médian, et les habenula, 4. au-dessus, le noyau caudé, 5. en avant, la tête du noyau caudé, et 6. en arrière, la queue du noyau caudé et le triangle habénulaire.
Il reçoit des afférences de l'ensemble du cortex cérébral, et forme le dernier relais des entrées sensorielles vers le cortex sensoriel primaire.
Histoire
Le thalamus est connu depuis longtemps puisque son nom vient de Galien (131-201 apr. J.-C.). Son nom complet était thalamus nervorum opticorum (ou thalamus opticus ou « couches optiques ») impliquant un lien avec les nerfs optiques. Le terme de pulvinar est de la même époque. Le thalamus a été ensuite inclus dans des ensembles confus (corpus striatum, Vieussens, « noyaux gris centraux », etc.). La connaissance de son lien avec le cortex est elle aussi ancienne. Mais le thalamus n'est ressorti comme entité particulière que depuis peu. Le concept d'isothalamus a simplifié considérablement la description en définissant deux entités séparés, l'isothalamus évitant de répéter l'analyse de la structure dans chacun de ses éléments.
Fonction
Le thalamus fonctionne comme une passerelle permettant à des informations sensorielles prétraitées d'entrer dans le néocortex. En particulier, la communication continue du thalamus avec le néocortex semble jouer un rôle préponderant. En effet, des dommages importants de la région principale du thalamus peuvent entraîner une perte de conscience prolongée. Un cas célèbre, celui de Karen Ann Quinlan, âgée de 21 ans, qui a subi une crise cardiaque et une insuffisance respiratoire et est restée dans un état végétatif apparemment sans réaction pendant dix ans. À sa mort, l'autopsie a révélé que son néocortex était normal mais que son thalamus avait été détruit[1].
Anatomie
Les thalamus sont des masses nerveuses obliques (environ 30° du plan ventriculaire médian chez l'humain, Percheron, 2003) et symétriques situées de part et d'autre du troisième ventricule. Ils sont parfois (dans 30 % des cas) liés entre eux par l'adhesio interthalamica mais dans notre espèce, celle-ci n'offre pas de connexion interthalamique.
L'intérieur du thalamus est séparé par des parties limitantes : des lamelles de substance blanche ou des ilots neuronaux particuliers. Depuis Burdach (1822) puis Dejerine (1901) et Foix et Nicolesco (1925), ces parties thalamiques sont utilisées pour faire un cloisonnement, distinguer et nommer les parties thalamiques. Certains noyaux thalamiques classiques (délimités par les éléments limitant ont été par la suite subdivisés. Le congrès de Louvain (voir Dewulf, 1971) a recommandé d'appeler « Régions » les noyaux classiques. Les nuclei en sont leurs parties ; pouvant avoir des pars. Une région peut être faite d'un ou de plusieurs noyaux. Parmi les « éléments limitants » neuronaux, il y a la substance grise périventriculaire, les éléments intralaminaires ou le nucleus limitans. Puisqu'ils n'ont pas la même constitution neuronale que celle de la majeure partie du thalamus, à la suite des Vogts pour le cortex, il a été proposé de les regrouper en un allothalamus opposé à l'isothalamus (Percheron, 2003). L'allothalamus est un ensemble hétérogène qui comprend la Formatio Paramediana (periventricular) et la Formatio Intralaminaris-limitans d'une part et la Regio Centralis qui correspond au « complexe centre médian-parafasciculaire » (qui a en fait trois subdivisions chez le primate supérieur) d'autre part. L'allothalamus, hétérogène, sera étudié après.
Isothalamus
Plus de 90 % du thalamus est en effet isothalamique. En dépit d'une grande variété de systèmes afférents, l'isothalamus est fait de peu d'éléments, arrangés de manière relativement simple (deux types de neurones) et utilisant peu de médiateurs (essentiellement deux : le glutamate et le GABA).
Les neurones constituants appartiennent à deux genera neuronaux hautement différenciés. Les premiers, les plus nombreux, correspondent au neurones thalamo-corticaux (ou principaux, Golgi I) qui sont très caractéristiques. On les dit « radiés » ou « touffus » car leur arborisation dendritique part de troncs épais et courts suivis de branches distales longues et raides. Le nombre de branches et le diamètre de l'arborisation dépend du système anatomo-fonctionnel considéré et de l'espèce animale mais pour chacun est fort stable, ce qui permet de définir des espèces neuronales. Les neurones thalamocorticaux ont la particularité de ne pas avoir de collatérales axonales initiales, ce qui implique qu'un neurone thalamocortical n'informe pas ses voisins de ce qu'il fait. De manière plus générale, il n'y a pas de connexion entre éléments isothalamiques. Les neurones thalamocorticaux ont des axones longs épais et myélinisés. Leur médiateur est le glutamate. L'autre genus neuronal est constitué de « microneurones » (Golgi II). Ils ont des branches dendritiques et axonales courtes. Ils contribuent à la circuiterie locale (interneurones). Leur pourcentage par rapport aux neurones thalamocorticaux dépend du système (à peu près 25 % dans les régions sensorimotrices) et de l'espèce animale, avec une forte augmentation chez les primates supérieurs. Leur axones courts ont pour cible les dendrites des neurones thalamocorticaux et d'autres microneurones. Leur médiateur est le GABA. Parmi les axones afférents sous-corticaux on doit distinguer ceux qui proviennent du périthalamus (noyau réticulé en particulier). Les neurones périthalamiques reçoivent des afférences (collatérales) des axones thalamo-corticaux. Il réémettent en retour des axones vers le cortex formant ainsi des circuits GABA inhibiteurs (circuits des Scheibel). Il y a dans l'évolution une évolution inverse entre les neurones perithalamiques qui déclinent et les microneurones dont le nombre s'accroit considérablement (Arcelli et al., 1997).
Région supérieure S
Cette région réunit deux éléments liés pendant longtemps et que l'on a, à tort, séparés plus tard : le noyau antérieur (nucleus anterior) et le noyau superficiel (nucleus superficialis) ou superior (inadéquatement appelé lateralis dorsalis). Le noyau antérieur, subdivisé dans les espèces non-humaines est indivis chez l'homme. Le noyau superficiel est situé en arrière de l'antérieur. Les deux noyaux, antérieur et superficiel, sont séparés des régions médiale et latérale par la lame supérieure. Ils sont partout entourés d'une capsule de matière blanche, incluant la lame terminale. L'ensemble est donc presque partout fermé. Les deux noyaux ont la même constitution. Ce qui les différencie est leur mode d'afférentation. Les deux reçoivent des informations du subiculum de l'hippocampe mais le noyau antérieur le fait de manière indirecte alors que le superficiel le fait directement. Les neurones efférents du subiculum forment le fornix qui passe au-dessus du thalamus pour terminer devant lui en avant du foramen de Monro. Là, une partie des axones descend pour rejoindre le corps mamillaire. Les neurones des corps mamillaires créent un faisceau épais et dense observé pour la première fois par Vicq d'Azyr (1783), le faisceau mamillo-thalamique (ou de Vicq d'Azyr) qui atteint le noyau antérieur par sa partie basse. Une autre partie des neurones du fornix ne se rendent pas au corps mamillaire mais tourne à la partie inférieure du foramen de Monro et se dirigent vers l'arrière. Ils fournissent un peu le noyau antérieur mais surtout le noyau superficiel. La cible corticale élective du noyau antérieur est le cortex cingulaire antérieur. Celle du noyau superficiel est le cortex cingulaire postérieur mais il y a chevauchement. De plus les deux aires cingulaires sont liées entre elles par le cingulum, important faisceau qui véhicule aussi les efférences cingulaires vers le cortex parahippocampique. Le « circuit de Papez » (1937) était déclaré par son auteur être le substrat de l'émotion. De nombreuses élaborations, incluant le « système limbique » ont suivi. En fait le circuit de Papez n'était pas fermé au niveau de l'hippocampe (et donc pas un circuit). Par ailleurs le second noyau, le noyau superficiel n'était pas pris en considération alors qu'il a des connexions similaires et participe à d'autres circuits. L'ensemble est multiple et complexe. L'effet le mieux établi de la lésion du corps mamillaire, du faisceau mamillothalamique et du fornix, si elle est bilatérale est une amnésie particulière, le syndrome de Korsakoff.
Région médiale. Noyau médial M
La région médiale, le nucleus medialis est le « noyau interne » de Dejerine) c’est-à-dire la région du thalamus située en dedans de la lame medullaire (lamina medialis). La partie antérieure de la lame qui contient des neurones intralaminaires constitue un bord clair. Ce n'est plus le cas en arrière où la séparation d'avec le pulvinar est loin d'être complète. Compte tenu de leur constitution et de leurs connexions similaires, les noyau medial et le pulvinar forment chez l'humain un ensemble qui occupe la majeure partie (en volume) du thalamus. Chez le singe on sépare dans le noyau médial habituellement trois parties parasagittales : de dedans en dehors une pars magnocellularis, une pars mediocellularis et une pars paralamellaris avec des efférences propres (Akert). Ceci ne s'observe pas chez l'homme qui n'a qu'un noyau indivis, le nucleus medialis. Celui-ci reçoit des afférences basses chez le singe : tectale et amygdalienne. Rien ne permet d'affirmer leur présence chez l'homme. L'essentiel des connexions est ainsi corticothalamique et thalamocortical. Les relations avec le cortex frontal sont connues depuis longtemps. Les lobotomies et tractotomies avaient pour but de les interrompre.
Région postérieure. Pulvinar. Pu
Le pulvinar (« coussin » en latin) représente la plus grande masse thalamique de l'humain. Il en est la partie la plus postérieure et constitue le pôle du thalamus. En avant, il n'a qu'une frontière partielle avec le noyau médial, et tous deux ont la même constitution et ont des connexions thalamo-corticales ou cortico-thalamiques communes. C'est le cas par exemple pour le cortex frontal. Les divisions faites dans le pulvinar sont abusives et n'ont pas la valeur de noyaux que peuvent avoir les noyaux latéraux. Elles ne correspondent pas à la distribution des afférences corticales. Une partie médiale ou principale reçoit des afférences du cortex frontal, pariétal, temporal et préoccipital. Une seule partie est particulière, le pulvinar intergeniculé ou inférieur qui reçoit chez le singe des afférences tectales (du colliculus supérieur) avec une carte visuotopique.
Région basale B
Dans la partie postéro-inférieure du thalamus, dans un lieu habituellement considéré comme situé dans le pulvinar, existe un endroit qui pose aujourd'hui des problèmes. Le faisceau spinothalamique termine dans trois entités différentes : intralaminaires-limitantes, « latérales » (dans le VCP, le VCO et le VI) et dans la formation basale (qui n'est pas un noyau classique) (C. Vogt, 1909, repris de Déjerine). Sa description a donné lieu récemment à des interprétations adverses. Une entité curieusement retenue comme VPMo (Blomquist, Craig, Zang) n'est pas ventrale et n'appartient pas au VP puisqu'elle ne reçoit pas d'afférences lemniscales, a été interprétée comme un centre particulier de réception des messages douloureux venant de la couche I de la moelle épinière. Il est observable (nucleus basalis nodalis), en particulier par la révélation de la calbindine et est entouré d'un halo. Cet endroit a été démontré envoyer des axones à l'insula. Il est par contre certain que d'autres axones nociceptifs se rendent aussi dans le noyau somesthèsique VPC. Cette partie du thalamus est l'objet de travaux et de controverses.
Région géniculée G
Le terme « métathalamus » qui regroupait les deux corps genouillés, n'est plus reconnu. Il était composé des deux corps genouillés (« en forme de genou », terme lié à la forme de l'externe) situés à la surface ventrale du thalamus, sous le pulvinar. Ils ont une face superficielle visible après dissection. Ce sont des « relais » de systèmes hautement différenciés : l'audition pour le médian et la vision pour le second. Ils se différencient très tôt dans l'ontogénèse au point qu'on les a classés longtemps hors thalamus. Le noyau géniculé latéral se sépare entièrement de la région latérale. le noyau médian reste attaché à la masse latérale. Quoique très spécialisés ce sont, tous deux, d'authentiques éléments isothalamiques.
Noyau géniculé médian auditif GM
Le nucleus geniculatus medialis reçoit des axones du système auditif. De la cochlée, l'information auditive périphérique se rend au noyau cochléaire. À partir de là, par le nerf cochléaire, les axones atteignent le complexe olivaire supérieur des deux côtés. De là, les axones forment le lemniscus latéral qui termine dans le colliculus inférieur. Les axones du colliculus inférieur forment le brachium du colliculus inférieur et terminent dans le noyau genouillé médian. Les axones thalamocorticaux du noyau géniculé médian terminent dans le cortex auditif primaire situé au centre de du plan temporal supérieur. Voir système auditif.
Noyau géniculé latéral GL
Le nucleus geniculatus lateralis est fait de plusieurs strates neuronales séparées par des lamelles parallèles à la surface mais fortement courbées. Les strates 1 et 2, les plus ventrales, sont magnocellulaires. Les autres sont médiocellulaires. À partir de la rétine, les axones des nerfs optiques vont directement aux noyaux geniculés. Un composant du nerf optique de chaque côté croisent au niveau du chiasma optique. Les axones rétiniens de la rétine controlatérale terminent dans les strates 1, 4 et 6. Ceux de la rétine ipsilatérale terminent dans les strates 2,3 et 5. Les axones du noyau géniculé latéral au sein des radiations optiques terminent dans le cortex visuel primaire, situé dans et autour de la fissure calcarine. Voir système visuel.
Région latérale L (ou V)
La région latérale correspond au « noyau externe » de Déjerine. Elle est subdivisée par l'arrivée (par le bas) de nombreux systèmes afférents. Les axones des systèmes importants s'emparent d'une portion de la masse latérale qu'ils ne partagent pas. Ainsi sont créés des « territoires » que l'on a rapproché des « noyaux ». Il n'y a pas d'opposition entre « noyaux ventraux » et « dorsaux ». Les éléments situés dorsalement sont en effet des parties postérieures de territoires plus antérieurs. Le choix du terme « ventral » plutôt que celui de « latéral » est plus d'usage que de logique anatomique. La cytoarchitectonique n'est pas toujours le moyen le plus fiable pour distinguer des sous-parties de la région latérale ou « noyaux ». L'immunocytochimie en particulier pour l'acetylcholinesterase ou pour les calcium binding proteins s'est révélée plus précise pour établir une subdivision structuro-fonctionnelle et pour faire des comparaisons entre espèces. Il y a des différences avec les non primates, mais même entre les singes et l'être humain.
Territoire gustatif VArc
Il est situé à la partie basse et antérieure du « noyau arqué » classique. Celui-ci n'est pas une entité simple. Sa partie supérieure est en effet un élément du territoire tactile VPM. Le noyau ventral arqué VArc à la partie inférieure du « noyau arqué » classique est à petites cellules et ne reçoit pas d'afférences lemniscales. Ce n'est donc pas une partie de VP. Il reçoit par contre des afférences gustatives du noyau du faisceau solitaire. Ses neurones thalamocorticaux envoient des axones à l'aire gustative primaire située dans l'opercule de l'insula (voir système gustatif).
Territoire lemniscal tactile VPC = VPM + VPL
Les noyaux correspondant au territoire lemniscal sont appelés VP (ventral postérieur). À l'intérieur de ce territoire, les cartes physiologiques (Mountcastle, Friedman et Jones), y compris chez l'homme (Albe-Fessard et al.), ont montré la séparation spatiale entre la sensibilité cutanée tactile et la sensibilité dite profonde (muscles, tendons, articulations). La représentation de la sensibilité tactile, plus en arrière, est précédée et surmontée par la représentation de la kinesthésie. Ceci est pris en compte dans la distinction formelle entre VPC (caudal) et VPO (oral). La partie tactile nucleus ventralis posterior caudal VPC est la partie postérieure de la région latérale, située en avant du pulvinar. C'est l'addition du noyau latéral VPL et de la partie supérieure du noyau arqué VPM. VPC reçoit les axones des noyaux des colonnes postérieures situés dans le bulbe : le nucleus gracilis (Goll) médial et le nucleus cuneatus (Burdach) latéral qui eux-mêmes reçoivent les axones du cordon postérieur de la moelle. À partir des noyaux postérieurs, les axones vont en avant et décussent (c'est-à-dire croisent pour changer de côté), toujours dans le bulbe (décussation lemniscale). Les axons des deux côtés forment l'épais lemnisque médian (lemniscus medialis) situé près de la ligne médiane. Celui-ci monte et devient latéral de chaque côté pour atteindre le bord inférieur du thalamus au niveau du VPC. Là, les axones terminent de manière dense en formant des lamelles en forme d'oignon. VPC forme un tout, avec une seule carte somatotopique. La représentation de la jambe est la plus postérieure, latérale et dorsale. Celle de la bouche et de la langue est la plus médiane, et la plus ventrale (dans VPM). Le médiateur est le glutamate. Les neurones thalamocorticaux du VPC envoient leurs axones à l'aire sensitive primaire constituée du koniocortex sensoriel (aire 3b) et du parakoniocortex (aire 1) où l'on observe aussi une somatotopie forte.
Territoire lemnical profond VPO (ou VPS)
À l'intérieur du territoire lemniscal, des cartes électrophysiologiques, y compris chez l'humain (Albe-Fessard), ont montré une séparation spatiale entre les représentations tactiles cutanées et « profondes » (ce qui réfère en général aux muscles, tendons et articulations). Chez le macaque, après Mountcastle, Friedman et Jones (1986) ont opposé la coque (« shell ») profonde d'un cœur (« core ») cutané. Kaas et al. (1986) après avoir retenu deux parties VPO et VPS sont revenus à une seule entité qu'ils ont appelée VPS. En fait la représentation de la sensibilité profonde forme une virgule à la fois au-dessus et en avant de la représentation cutanée. Dans la séquence postéro-antérieure des territoires, la représentation profonde se trouve ainsi entre le VPC tactile et le territoire cérébelleux (VIm), d'où la préférence pour le sigle VPO opposé au VPC tactile. (Le VPO est l'addition des VPO et VPS de Kaas). VPO est constitué de neurones particulièrement grands (les plus grands du thalamus) avec des amplitudes de pointes particulièrement élevées. Ces neurones reçoivent leurs afférences du nucleus nucleatus accessorius du bulbe. Les axones de ce noyau décussent et participent à la formation du lemniscus médian où ils restent latéraux. Ils atteignent la partie inférieure du thalamus pour emplir le territoire. Le VPO qui reçoit donc les informations musculaires, tendineuses et articulaires a une carte somatotopique presque identique à celle du VPC cutané. Les neurones du VPO ont une cible corticale élective, l'aire fundique 3a, dans la profondeur du sillon de Rolando (entre l'aire motrice 4 et l'aire somesthésique tactile 3b et 1) mais aussi l'aire pariétale 5.
Territoire cérébelleux VIm ou VL
Le médiateur du système cérebello-thalamique est le glutamate, excitateur. Le territoire cérébelleux reçoit des axones de tous les noyaux cérébelleux mais en particulier du noyau dentelé (Percheron, 1977 ; Asanuma et al., 1983). Il existait déjà des arguments en faveur d'une subdivision du territoire cérébelleux mais une découverte marquante récente est que le noyau dentelé, chez le primate, est subdivisé en deux parties fonctionnelles : une antérieure (« motrice ») et l'autre postéroventrale non motrice, « associative » (Dum and Strick, 2002). Cette subdivision induit sans doute celle du territoire cérébelleux. Le nucleus ventralis intermedius est en fait composé de deux parties, une ventrolatérale (VImL) et une médiodorsale (VImM). La première est bien connue. VImL (le VIm des neurochirurgiens) reçoit électivement des informations sensorimotrices. Il reçoit aussi des axones du vestibule et du faisceau spinothalamique. Les afférences cérébelleuses y sont organisées selon une carte somatotopique qui reproduit grossièrement celle du VPC. La cible corticale élective de VImL est le cortex moteur primaire (prerolandique) (Schell and Strick, 1984, Orioli and Strick, 1989). VImM était décomposée en une partie médiale (l'« area X », V.im.i) et une partie dorsale (VLc) qui passe au-dessus de VImL et de VCL pour atteindre le bord dorsal du thalamus. Cet ensemble reçoit essentiellement des informations « associatives » du noyau dentelé postéroventral. Il reçoit aussi des axons tectaux et spinothalamiques. Il est organisé selon une autre seule carte, plus lâche, moins fine que celle de VImL. Les neurones thalamocorticaux de VImM envoient leurs axones au cortex prémoteur et pariétal. Comme il n'avait pas été jusque-là sérieusement pris en compte, il n'y a que peu de données physiologiques à son propos.
Territoire pallidal VO
À partir des cercopithecidae, les deux sources d'axones du système des ganglions de la base projetant au thalamus forment deux territoires distincts. Ceci explique en partie les problèmes rencontrés historiquement puisque ce n'est pas le cas des espèces inférieures. Le pallidum median et la nigra ont chacun un territoire propre. Le territoire pallidal apparait quand un pallidum médian important envoie des axones à la partie latérale du thalamus, latéralement au territoire nigral VA. C'est donc un nouveau noyau qui s'ajoute chez le primate, le nucleus ventralis oralis, VO. Puisqu'il n'y a plus de territoire commun comme chez les rongeurs et les carnivores, il n'y a plus de VM qui est topographiquement une partie de VA. VO reçoit ses axones pallidaux du pallidum médian. Leur trajectoire est spatialement très complexe. Les axones forment d'abord l'ansa lenticularis et le fasciculus lenticularis qui placent les axones au bord médial du pallidum. À partir de là, les axones pallidaux traversent la capsule interne comme système en peigne d'Edinger. Les axones arrivent au bord latéral du nucleus subthalamique de Luys Ils passent au-dessus de lui comme champ H2 de Forel (1877) et tournent en descendant en H. Là, soudainement ils montent vers le bord inférieur du thalamus (VO) formant H1. La distribution des axones à l'intérieur du territoire est lâche. Un même axone pouvant couvrir un espace important. Les extrémités émettent des « bouquets » denses (Arrechi-Bouchhioua et al., 1996, 1997 ; Parent and Parent, 2004). Ceci n'offre qu'une chance faible à une organisation somatotopique fine. Le médiateur de la connexion pallido-thalamique est le GABA inhibiteur. Les neurones thalamocorticaux du VO ont pour cibles principales l'aire motrice supplémentaire (SMA), la preSMA et à un degré moindre le cortex moteur primaire.
Territoire nigral VA
VA a longtemps eu un statut incertain puisqu'on allait jusqu’à dire qu'il n'avait pas de projection corticale! Ni ses limites ni ses fonctions n'étaient clairement établies. Il est important de lui restituer sa place dans le thalamus moteur en le distinguant du territoire pallidal. VA est en effet le territoire nigral. Les axones afférents viennent de la pars reticulata de la noire. Les axones nigrothalamiques ne constituent pas un faisceau évident. Ils forment une nappe parasagittale située en dedans des afférences pallidales qui monte verticalement. Dans le thalamus, ils forment VA qui inclut le faisceau mamillaire de Vicq d'Azyr. Une partie du territoire, soit négligée soit faussement attribuée à un VM, est postérieure et ventrale, étendue jusqu'au pôle de la région centrale. Les deux territoires pallidaux et nigraux étant partout séparés chez le primate supérieur, il n'y a plus de VM. Dans le VA, les arborisations axonales ont une distribution très large (François et al., 2002) ne permettant pas de carte précise. Le médiateur de la connexion nigro-thalamique est GABAergique inhibitrice. En plus de l'afférence nigrale, VA reçoit des axones amygdaliens et tectaux (colliculus supérieur). Les axones thalamocorticaux se rendent au cortex frontal, au cortex cingulaire et au cortex ocilomoteur FEF et SEF. Ceci donne une indication sur le caractère visuomoteur du noyau dont la physiologie reste mal connue.
Éléments allothalamiques
Les éléments allothalamiques sont hétérogènes. On peut distinguer deux sous-groupes : le noyau circularis (ou involucrum) composé des éléments paramédians et intralaminaires et la région centrale.
Formation paramédiane
Chez l'humain, elle est limitée à une nappe neuronale mince située juste latéralement à l'épendyme. La division en noyaux n'a pas lieu d'être. Cet ensemble est lié aux systèmes périventriculaires.
Formation intralaminaire limitante Il-LI
La formation intralaminaire est réduite aux éléments intralaminaires (à l'intérieur de la lamina medialis) situés en dehors du noyau médial qu'elle entoure, auxquels s'ajoutent les neurones des groupes en arrière de la région centrale (non décrits chez le singe) et du « noyau limitans » de Friedemann (1911) étendu sous le pulvinar qu'il sépare du mésencéphale. Les éléments postérieurs, y compris le limitans, reçoivent des axones spinothalamiques. La présence d'axones cérébelleux, et tectaux, certaine chez le singe, n'est pas attestée chez l'humain. Les axones de cette formation envoient des axones au striatum. Ils prennent une part relativement faible à la connexion thalamo-striée par rapport à la région centrale (voir plus loin).
Région centrale C
Elle correspond au « complexe centre médian-parafasciculaire ». Le « centre médian » est le noyau décrit par Luys, 1865. Le noyau parafasciculaire a été décrit plus tard, chez le singe, par Friedemann (1911). En fait, chez les primates supérieurs, à partir des cercopithecidae, la région n'est pas constituée de deux mais de trois parties, chacune avec sa propre espèce neuronale et des connexions particulières (Fénelon et al., 1991). Contrairement à ce qui est généralement dit, cet ensemble neuronal n'appartient pas au groupe des « noyaux intralaminaires ». Ceci s'affirme sur son ontogénèse, sa position, sa structure et ses connexions. C'est un ensemble bien caractérisé et très évolutif. Il est presque partout entouré d'une capsule. Dorsalement celle-ci est constituée du prolongement ventral de la lamina medialis (lame médullaire interne). Latéralement la région est séparée de la région latérale par la lamina centralis. La présence de trois parties a entrainé un débat sur l'appartenance de la partie intermédiaire au parafasciculaire (Niimi et al., 1960) ou au centre médian (les Vogt, 1941). C'est en fait indécidable. On décrit donc une région centrale (qui est un noyau classique) avec de dedans en dehors une pars parafascicularis, une pars media et une pars paralateralis. Les deux premières se colorent fortement pour l'acetylcholinesterase. Elles ont des connexions fortes avec le système des ganglions de la base. La pars parafascicularis est liée bilatéralement à la nigra reticulata. Elle reçoit par ailleurs des axones du colliculus supérieur. Elle envoie des axones au striatum associatif. Elle fait partie d'un circuit striatum-nigra reticulata-pars parafascicularis- striatum probablement lié à des tâches visuomotrices. La pars media elle, reçoit des axones du pallidum médian. Elle reçoit aussi des afférences du cortex moteur et moteur accessoire. Elle envoie ses neurones au striatum sensorimoteur (Fenelon et al., 1991). Elle est un élément du circuit de Nauta-Mehler (striatum-pallidum-pars media-striatum). Les deux éléments médians sont les premiers contributeurs de la connexion « thalamo-striée ». Leur médiateur est le glutamate. Le troisième élément le plus latéral n'apparait qu'avec les primates supérieurs (à partir des cercopithecidae) où il prend un développement important, la pars paralateralis. Celle-ci reçoit et envoie des axones du cortex moteur et prémoteur. La région centrale n'est donc pas une part d'un système thalamique nonspécifique. C'est un élément important du système des ganglions de la base dont il est un des régulateurs (voir système des ganglions de la base du primate Primate Basal Ganglia system).
Notes et références
- (en) Ray Kurzweil, How to Create a Mind : The Secret of Human Thought Revealed, Prelude Books, , 336 p. (ISBN 978-0-7156-4595-6, lire en ligne)
- Arcelli P, Frassoni C, Regondi M, De Biasi S, Spreafico R (1997) GABAergic neurons in mammalian thalamus: a marker of thalamic complexity? Brain Res Bull 42 :27-37.
- Arecchi-Bouchhioua P, Yelnik J, Francois C, Percheron G, Tande D.(1996) 3-D tracing of biocytin-labelled pallido-thalamic axons in the monkey. Neuroreport.7:981-984.
- Arrechi-Bouchhioua, P., Yelnik, J., Percheron, G., Tande, D. (1997) Three dimensional morphology and distribution of pallidal axons projecting to both the lateral region of the thalamus and the central complex in primate. Brain Res. 754:311-314.
- Asanuma, C., Thach, W.T. and Jones, E.G. (1983) Anatomical evidence for segregated focal groupings of efferent cells and their terminal ramifications in the cerebellothalamic pathway in the monkey. Brain Res. Rev. 5: 267-297
- Attempt at standardization of nomenclature. In Dewulf, A. (1971) Anatomy of the normal human thalamus. Topometry and standardized nomenclature. Elsevier, Amsterdam p. 121-139
- Burdach, K. F. (1822) Von Baue und Leben des Gehirns. Dyk, Leipzig
- Dejerine, J. and Roussy. G.(1906) Le syndrome thalamique. Rev. Neurol. (Paris)14: 521-532
- Dum, R.P., Li, C., and Strick., P.L.(2002) Motor and non motor domains in the monkey dentate.Ann. N.Y.Acad. Sci. 978:289-301.
- Fenelon, G., François, C., Percheron, G. and Yelnik, J. (1991) Topographic distribution of neurons of the central complex (centre médian-parafascicular complex) and of other thalamic neurons projecting to the striatum in macaques. Neurosci.45:495-510
- Fenelon, G., Yelnik, J., François, C. and Percheron, G. (1994) The central complex of the primate thalamus: a quantitative analysis of neuronal morphology. J. Comp. Neurol. 341: 463-479
- François, C., Tande, D., Yelnik, J., and Hirsch, E.(2002) Distribution and morphology of nigral axons projecting to the thalamus in primates. J. Comp. Neurol. 447:249-260.
- Friedman, D.P. and Jones, E.G. (1986) Thalamic input to area 3a and 2 in monkeys. J. Neurophysiol. 45: 59:85
- Goldman-Rakic, P.S. and Porrino, L.J. (1985) The primate dorsomedial (MD) nucleus and its projection to the frontal lobe. J. Comp. Neurol. 242:535-560
- (en) Orioli, P.J. and Strick, P.L., « Cerebellar connectionswith the motor cortex and the arcuate premotor area:an analysisemploying retrograde transneuronal transport of WGA-HRP. », J. Comp. Neurol., no 288, , p. 612-626 (PMID 2478593)
- Kaas, J.H., Nelson, R.J., Sur, M. Dykes, R.W., Merzenich, M.M (1984) The somatotopic organisation of the ventroposterior thalamus of the squirrel monkey, Saimiri sciureus. J. Comp. Neurol. 226:111-140
- Papez, J.W. (1937) A proposed mechanism of emotion. Arch. Neurol. Psychiat.38:725-743.
- Parent, M. and Parent, A. (2004) The pallidofugal motor fiber motor system in primates. Park. Relat. Disord. 10: 203-211
- Percheron, G. (1977) The thalamic territory of cerebellar afferents in macaques . J.Hirnforsch. 18: 375-400
- Percheron, G. (2003) Thalamus. In Paxinos, G. and May, J.(eds). The human nervous system. 2d Ed. Elsevier. Amsterdam. p. 592-675
- Percheron, G., François, C., Talbi, B., Meder, J.F, Fenelon, G.and Yelnik, J. (1993) The primate motor thalamus analysed with reference to subcortical afferent territories. Stereotact. Funct. Neurosurg. 60: 32-41
- Percheron, G., François, C., Talbi, B., Meder, J.F, Yelnik, J, and Fenelon, G.(1996) The primate motor thalamus. Brain Res. Rev. 22:93-181
- Percheron, G., François, C., Yelnik, J, Talbi, B., Meder, J.F and Fenelon, G.(1993) The pallidal and nigral thalamic territories and the problem of the anterior part of the lateral region in primates. In Minciacchi., D. et al. (eds) Thalamic networks for relay and modulation. Pergamon Press.Oxford p. 145-154
- (en) Rockland K, « Further evidence for two types of corticopulvinar neurons », Neuroreport, vol. 5, no 15, , p. 1865-8 (PMID 7841364)
- (en) Schell, E.R. and Strick, P.L., « The origin of thalamic inputs to the arcuate premotor and supplementary motor areas. », J. Neurosci., no 4, , p. 539-560 (PMID 6199485)
- Terminologia anatomica (1998) Thieme, Stuttgart. (ISBN 3131143614)
- Vogt, C. (1909) La myelocytoarchitecture du thalamus du cercopithèque. J. Psychol. Neurol. 12: 285-324.