Teleocrater
Teleocrater rhadinus

de Teleocrater rhadinus.
Teleocrater est un genre éteint de « reptiles » primitifs archosaures du clade également éteint des aphanosaures. Il a vécu au cours l'âge anisien du Trias moyen en Tanzanie[1], soit il y a environ entre 247 et 242 Ma (millions d'années). Teleocrater est ainsi un des plus anciens archosaures avémétatarsaliens connus, si ce n'est le plus ancien avec Asilisaurus[2] - [1].
L'espèce type et la seule espèce rattachée au genre est Teleocrater rhadinus.
Les analyses phylogénétiques réalisées à la suite de la description de cette nouvelle espèce ont permis l'érection d'un nouveau clade, les Aphanosauria, dans lequel Nesbitt et ses collègues ont placé, en compagnie de Teleocrater, plusieurs genres d'archosaures dont la position taxonomique était jusqu'à présent mal connue et très discutée. Il s'agit des genres : Spondylosoma, Yarasuchus et Dongusuchus[1]. Les Aphanosauria se situent en groupe frère des ornithodires, un clade qui contient les dinosaures et les ptérosaures.
Description
Le genre a été nommé et décrit formellement en 2017 par Sterling Nesbitt et son équipe internationale de dix paléontologues à partir de fossiles trouvés dans un lit d'ossements de la formation géologique de Manda du Trias moyen de Tanzanie.
Teleocrater est un quadrupède carnivore à long cou, d'une longueur totale estimée entre 1,80 et 2,50 mètres. Il est remarquable par la longueur et la morphologie particulière de ses vertèbres cervicales[1]. À la différence des Lagerpetidae ou des ornithodires, ses membres postérieurs ne sont pas adaptés pour la course et ses métatarses ne sont pas particulièrement longs. De même, à la différence de ces deux groupes, Teleocrater a conservé la configuration plus flexible de la cheville, comme celle des archosaures primitifs suggérant que cette configuration était également présente chez les ancêtres des Avemetatarsalia mais qu'elle a été ensuite perdue indépendamment dans plusieurs lignées. Les études histologiques des os longs de l'animal indiquent que le taux de croissance de l'animal était relativement rapide plus proche de celui des ornithodires que de celui des crocodiliens ou des autres pseudosuchiens[1].
Crâne
La seule dent retrouvée sur le crâne de Teleocrater est comprimée, recourbée et porte des dentelures, comme des dents de scie, sur ses deux tranchants ; elle indique clairement un régime alimentaire carnivore. Comme les dinosaures primitifs, son os frontal montre une dépression en face de la fenêtre supra-temporale, formant une fosse supra-temporale[3] - [4] - [5] - [1].
Colonne vertébrale
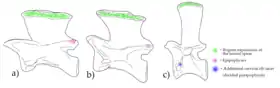
en vert le surplomb vers l'avant (vers la gauche) de l'épine neurale en forme de lame de hachette,
en rose des processus pointus (épipophyses),
et en bleu des facettes d'insertion de côtes cervicales.
Cervicales
Les vertèbres cervicales de la moitié antérieure du cou de l'animal sont assez longues, 3,5 fois plus hautes que larges. Elles figurent parmi les plus longues connues dans le clade des Avemetatarsalia. Elles portent des épines neurales en forme de lame de hachette avec des bords avant qui se rétrécissent en pointe et surplombent nettement le corps vertébral ou centrum de la vertèbre[6]. Elles porteant aussi d'autres processus arrondis et rugueux.
Les vertèbres cervicales de l'avant et du milieu du cou montrent des processus pointant vers l'arrière (épipophyses) ce qui a déjà été observé chez Yarasuchus[7] et chez certains pseudosuchiens[5] - [8] - [1].
Les vertèbres cervicales de la moitié arrière du cou montrent d'autres processus supplémentaires ; elles semblent avoir permis l'attache de côtes cervicales à trois têtes [6].
Dorsales
Les vertèbres dorsales sont caractérisées, comme chez tous les aphanosaures par la présence de petites dépressions sur le côté de leur base. Deux vertèbres sont associées au sacrum chez Teleocrater, un nombre proche des trois vertèbres liées au sacrum du genre de dinosauriformes Nyasasaurus qui a été découvert dans la même formation géologique tanzanienne que Teleocrater[9].
Les côtes associées aux dernières vertèbres sacrées montrent des processus pointés vers l'arrière ou le côté, ce qui n'est connu que chez Yarasuchus et Spondylosoma et les dinosauriformes. Aucun ostéoderme n'a été découvert avec les squelettes de Teleocrater, ce qui le différencie des pseudosuchiens[1].
Pattes avant
Comme les autres archosaures et les membres de la famille des protérosuchidés[8], Teleocrater possède un acromion, un processus distinct, sur son omoplate et, comme chez les silésauridés, celui-ci porte une étroite crête sur sa partie arrière.
Son humérus présente une longue crête delto-pectorale qui s'étend sur environ 30% de la longueur de l'os ; une caractéristique partagée avec les autres aphanosaures mais aussi chez le dinosauriforme Nyasasaurus découvert dans la même formation géologique[10] - [9]. Cette crête n'existe pas chez les ptérosaures et les silésauridés. Une autre caractéristique des aphanosaures, que possède Teleocrater, est une extrémité distale de l'humérus très large (environ 30% de la longueur de l'os). Ses mains sont de taille assez modeste[1].
Pattes arrière
Le nom de Teleocrater indique que sa cavité cotyloïde de l'os iliaque, où se loge la tête du fémur, est quasiment fermée. Il y a une petite encoche concave sur le bord inférieur de la partie de l'os iliaque qui s'étend jusqu'à rencontrer l'ischium, ce qui suggère la présence d'une petite perforation à l'intérieur de la cavité cotyloïde. Cette morphologie est connue également chez les silésauridés Asilisaurus[2] et Silesaurus[11].
Comme chez Asilisaurus[2] et Marasuchus, un Dinosauriformes[12], la partie antérieure de l'os iliaque est séparée du reste de l'os par une crête qui s'élève verticalement à partir du bord supérieur de la cavité cotyloïde[1].
Les proportions des différents os des pattes arrière de Teleocrater rappellent celles des silésauridés, des pseudosuchiens et des archosaures primitifs, avec des métatarses pas particulièrement longs par rapport au fémur et au tibia. L'allongement relatif des métatarses chez les groupes qui viendront plus tard représente probablement une adaptation à la course[13] - [1].
Le fémur de l'animal montre une combinaison de caractéristiques variées. Comme pour les autres aphanosaures, la tête du fémur exhibe un sillon transversal, tandis que son épiphyse distale montre une surface concave[1]. Le fémur de Teleocrater rappelle celui de son très proche parent Dongusuchus, avec cependant, chez Teleocrater, des côtés de la tête de fémur plus arrondis et une surface de contact plus concave[1].
Contrairement aux proterochampsidés ou aux dinosaures, la face avant de la tête du tibia de Teleocrater ne porte pas de crête cnémiale (qui sert à l'attachement du muscle extenseur de la cuisse)[8] - [1]. Le péroné présente une longue crête torsadée et le bord antérieur du sommet de l'os est élargi vers l'extérieur. Le calcanéum de Teleocrater et des aphanosaures en général, mais aussi des silésauridés (à savoir Asilisaurus et Lewisuchus[2]), et des pseudosuchiens, sont similaires. Ils présentent tous une articulation convexe-concave avec l'astragale qui permet le libre mouvement, une différence avec les dinosaures[1].
Paléoécologie
Le lit d'ossements ayant fourni les fossiles de Teleocrater a également livré deux populations d'os :
- de grands os de dicynodontes du genre Dolichuranus sp. et de cynodontes du genre Cynognathus sp.[14]. Ces os de grande taille sont accumulés, parfois encore semi-articulés, indiquant un transport limité par l'eau après la mort des animaux ;
- de petits os appartenant à Teleocrater, au temnospondyle "Stanocephalosaurus" pronus, à un allokatosaure inconnu et à un autre reptile de petite taille non nommé. Ces os sont souvent fragmentés, ce qui suggère qu'ils ont été transportés sur de plus longues distances ou plusieurs fois avant de se sédimenter dans la plaine alluviale[15] - [16]. D'autres niveaux stratigraphiques de la formation de Manda ont fourni des fossiles du ctenosauriscidé Hypselorhachis mirabilis[17] - [1].
Classification
Sterling Nesbitt et une équipe internationale de dix paléontologues définissent en 2017 le clade des Aphanosauria, comme le plus inclusif contenant Teleocrater rhadinus et Yarasuchus deccanensis, mais pas Passer domesticus ou Crocodylus niloticus. Leur cladogramme ci-dessous montre la position de Teleocrater dans une polytomie, incluant aussi les genres Dongusuchus et Yarasuchus, en groupe frère du genre brésilien Spondylosoma[1] :
Notes et références
- (en) S.J. Nesbitt, R.J. Butler, M.D. Ezcurra, P.M. Barrett, M.R. Stocker, K.D. Angielczyk, R.M H. Smith, C.A. Sidor, G. Niedźwiedzki, A.G. Sennikov et A.J. Charig, « The earliest bird-line archosaurs and the assembly of the dinosaur body plan », Nature, (DOI 10.1038/nature22037, lire en ligne)
- (en) S.J. Nesbitt, C.A. Sidor, R.B. Irmis, K.D. Angielczyk, R.M.H. Smith et L.A. Tsuji, « Ecologically distinct dinosaurian sister group shows early diversification of Ornithodira », Nature, vol. 464, , p. 95–98 (DOI 10.1038/nature08718, lire en ligne)
- (en) Stephen L. Brusatte, S. J. Nesbitt, R. B. Irmis, R. J. Butler, M. J. Benton et Mark A. Norell, « The origin and early radiation of dinosaurs », Earth-Science Reviews, vol. 101, no 1, , p. 68–100 (DOI 10.1016/j.earscirev.2010.04.001, lire en ligne)
- (en) M.C. Langer, M.D. Ezcurra, J.S. Bittencourt et F.E. Novas, « The origin and early evolution of dinosaurs », Biological Reviews, vol. 85, no 1, , p. 55–110 (DOI 10.1111/j.1469-185X.2009.00094.x, lire en ligne)
- (en) S.J. Nesbitt, « The early evolution of archosaurs: relationships and the origin of major clades », Bulletin of the American Museum of Natural History, vol. 352, , p. 1–292 (DOI 10.1206/352.1, lire en ligne)
- (en) Sterling J. Nesbitt, Richard J. Butler, Martin D. Ezcurra, Alan J. Charig et Paul M. Barrett, « The anatomy of Teleocrater Rhadinus, an early avemetatarsalian from the lower portion of the Lifua Member of the Manda Beds (Middle Triassic) », Journal of Vertebrate Paleontology, vol. 37, no sup1, , p. 142–177 (DOI 10.1080/02724634.2017.1396539, lire en ligne)
- (en) Kasturi Sen, « A new rauisuchian archosaur from the Middle Triassic of India. », Palaeontology, vol. 48, no 1, , p. 185–196 (DOI 10.1111/j.1475-4983.2004.00438.x, lire en ligne)
- (en) M.D. Ezcurra, « The phylogenetic relationships of basal archosauromorphs, with an emphasis on the systematics of proterosuchian archosauriforms », PeerJ, vol. 4, , e1778 (DOI 10.7717/peerj.1778, lire en ligne)
- (en) S.J. Nesbitt, P.M. Barrett, S. Werning, C.A. Sidor et A.J. Charig, « The oldest dinosaur? A Middle Triassic dinosauriform from Tanzania », Biology Letters, vol. 9, no 1, (DOI 10.1098/rsbl.2012.0949, lire en ligne)
- (en) M.C. Langer et M.J. Benton, « Early dinosaurs: A phylogenetic study », Journal of Systematic Palaeontology, vol. 4, no 4, , p. 309–358 (DOI 10.1017/S1477201906001970, lire en ligne)
- (en) J. Dzik, « A beaked herbivorous archosaur with dinosaur affinities from the early Late Triassic of Poland », Journal of Vertebrate Paleontology, vol. 23, no 3, , p. 556–574 (DOI 10.1671/A1097, lire en ligne)
- (en) P.C. Sereno et A.B. Arcucci, « Dinosaurian precursors from the Middle Triassic of Argentina: Marasuchus lilloensis, gen. nov. », Journal of Vertebrate Paleontology, vol. 14, no 1, , p. 53–73 (DOI 10.1080/02724634.1994.10011538, lire en ligne)
- (en) T. Kubo et M.O. Kubo, « Associated evolution of bipedality and cursoriality among Triassic archosaurs: a phylogenetically controlled evaluation », Paleobiology, vol. 38, no 3, , p. 474–485 (DOI 10.1666/11015.1, lire en ligne)
- (en) B.M. Wynd, C.A. Sidor, B.R. Peecook, M. Whitney, R.M.H. Smith, S.J. Nesbitt, K.D. Angielczyk et N.J. Tabor, First occurrence of Cynognathus in Tanzania and Zambia, with biostratigraphic implications for the age of Triassic strata in southern Pangea, vol. 36, Salt Lake City, (lire en ligne), p. 254
- (en) N.D. Smith, T.A. Cross, J.P. Dufficy et S.R. Clough, « Anatomy of an avulsion », Sedimentology, vol. 36, no 1, , p. 1–23 (DOI 10.1111/j.1365-3091.1989.tb00817.x, lire en ligne)
- (en) R.M.H. Smith, « Sedimentology and ichnology of floodplain paleosurfaces in the Beaufort Group (Late Permian), Karoo Sequence, South Africa », PALAIOS, vol. 8, no 4, , p. 339–357 (DOI 10.2307/3515265, lire en ligne)
- (en) R.J. Butler, P.M. Barrett, R.L. Abel et D.J. Gower, « A Possible Ctenosauriscid Archosaur from the Middle Triassic Manda Beds of Tanzania », Journal of Vertebrate Paleontology, vol. 29, no 4, , p. 1022–1031 (DOI 10.1671/039.029.0404, lire en ligne)
Voir aussi
Références taxinomiques
(en) Référence Paleobiology Database : Teleocrater Nesbitt et al., 2017 (en) Référence Paleobiology Database : Teleocrater rhadinus Nesbitt et al., 2017