Réponses plastiques et évolutives à la compétition intraspécifique
Il existe au sein d'une même espèce, animale ou végétale, des interactions entre les individus, notamment la compétition pour les ressources (alimentaires, reproductives, territoriales, etc.) cette compétition est appelée compétition intraspécifique.
Cette compétition intraspécifique peut prendre différentes formes par rapport à l'acquisition de cette ressource. Cependant, si une modification apparaît dans l'environnement telle qu'une compétition intraspécifique plus intense, un changement ponctuel ou permanent des conditions climatiques… L'espèce développe des réponses pour s'adapter à ses changements. Deux grands types de réponses sont distingués : la plasticité et l'évolution.
La plasticité est une réponse réversible ayant une influence sur le comportement d’un ensemble d’individu à un instant donné et dans des conditions données, qui ne sera pas transmis à la descendance.
L’évolution est une réponse irréversible jouant sur la sélection d’un caractère génétique procurant un avantage dans le milieu considéré à un individu et se transmettant via la reproduction.
Bien que se déroulant sur des échelles différentes, ces deux réponses coexistent au sein d'une même population. En effet, elles ne sont pas développées par tous les individus de l’espèce au même moment de leur cycle de vie, elles ne répondent pas aux mêmes contraintes.
La compétition et la compétition intraspécifique
Définition de la compétition
La compétition, au sens écologique du terme, désigne le combat entre les organismes pour l'accès à des ressources en quantité ou en qualité. Elle agit alors de manière négative sur la fitness ou, en français, valeur sélective de chaque individu de la population prise en compte. La fitness peut être exprimée par le nombre de descendant d'un individu ayant atteint la maturité sexuelle.
Les ressources peuvent être de différentes natures comme la nourriture, l’abri (choix de la localisation ou des ressources nécessaires à sa fabrication) ou encore l'accès à un partenaire sexuel pour assurer la reproduction. Elles constituent ce que l'on appelle la niche écologique de l'espèce considérée. Cette niche est définie par la quantité et la variété de ressources et d'abris que l'espèce peut exploiter. On distingue deux types de niches : la niche fondamentale et la niche réalisée.
La niche fondamentale est la niche que peut occuper l'espèce en l'absence de compétiteurs. Il s'agit d'une niche dite théorique.
La niche réalisée est la niche que peut réellement occuper l'espèce en présence de compétiteurs, l'empêchant ainsi d'occuper une partie de sa niche fondamentale.
On distingue deux formes de compétition : la compétition interspécifique qui est la compétition entre des individus d'espèces différentes et la compétition intraspécifique qui est la compétition entre individus de la même espèce.
La compétition intraspécifique
D'après De l'origine des espèces de Charles Darwin, la compétition intraspécifique est toujours plus importante que la compétition interspécifique[1]. Cela s'explique par le fait que les individus d'une même espèce sont exposés aux mêmes dangers et ont les mêmes besoins en termes de nourriture et d'abris.
Cette compétition peut prendre plusieurs formes.
La compétition par mêlée
Voir aussi Scramble competition
La compétition par mêlée (ou Scramble competition en anglais), est une compétition symétrique. C'est-à-dire, tous les compétiteurs pris en compte s'affectent de la même façon les uns les autres au niveau de l'accès aux ressources. Tous n'auront donc pas la quantité de ressources nécessaire à leur développement optimal. Si la ressource disparaît ou vient à manquer, tous les individus de la population en subiront les conséquences, pouvant à terme mourir[2].
Ce type de compétition est observable dans la nature :
- chez les végétaux, où la récupération des minéraux dans le sol par les racines s'effectue en fonction des ressources restantes laissées par les autres individus ;
- chez les animaux, où dans le cas des herbivores (tels les moutons ou encore les vaches), la quantité d'herbes restantes dans le pâturage va avoir un impact sur l'ensemble de la population.
Ce type de compétition a été étudié de manière plus détaillée par Jordi Moya-Laraño, Maysaa El Tigani El-Sayyid et Charles W Fox dans le cadre de l'observation de l'impact de températures fraîches sur la taille des scarabées mâles dans le cadre de la compétition pour la reproduction[3].
La compétition par concours
Voir aussi Contest competition
La compétition par concours (ou Contest competition en anglais), signifie que l'acquisition de la majorité des ressources sera faite par l'individu le plus fort[4]. L'individu le plus fort pouvant être caractérisé de différentes façons :
- le plus rapide, comme le vol chez les scarabées pour l'accès à la reproduction ;
- le plus voyant, comme la queue du paon là encore pour accéder à la reproduction ;
- le plus fort physiquement, comme pour la défense de la proie contre les charognards.
Certaines de ces caractéristiques peuvent s'additionner. D'autres éléments peuvent aussi entrer en compte.
La sélection sexuelle
La sélection sexuelle est un type de compétition par concours où la compétition se fait entre individus du même sexe pour l'accès à la reproduction.
Le sexe qui est en compétition est le sexe qui investit le moins d'énergie dans la reproduction. En général, il s'agit du mâle car les femelles ont des gamètes couteux à produire et ne peuvent avoir qu'une (ou qu'un petit nombre) portée ou ponte par saison de reproduction, contrairement aux mâles. Elles ont donc intérêt à choisir les mâles avec les meilleurs gènes pour assurer une meilleure survie à leur progéniture.
Selon les espèces, les mâles vont se battre pour l'accès aux femelles (sélection intrasexuelle) où vont être choisis par ces dernières sur des critères de physique, de qualité d'offrandes ou encore de taille de territoire (sélection intersexuelle).
Ces techniques utilisées par les mâles pour s'approprier les femelles sont en fait des moyens que les femelles ont de pouvoir estimer la qualité d'un mâle.
Les mâles les plus forts vont gagner les combats et donneront naissance à des enfants forts à leur tour ce qui augmentera la valeur sélective de la femelle.
Les mâles les plus «beaux» ont souvent un handicap lié à cette beauté, comme la queue du paon par exemple, ce qui témoigne de leurs bons gènes qui les aident à survivre malgré ce handicap (voir l'article détaillé Théorie du handicap).
Les mâles qui apporteront le plus de nourriture de bonne qualité à la mère permettront à cette dernière d'augmenter ses chances de survie et lui assure que ce mâle pourra apporter beaucoup de nourriture à sa progéniture.
La compétition fréquence-dépendante
Ce type de compétition apparaît lorsque dans une population certains individus sont aptes à exploiter une ou plusieurs ressources différentes de la niche écologique de l'espèce. Ainsi lorsque la ressource principale de l'espèce devient rare ou qu'il y a trop d'individus l'exploitant, ceux pouvant utiliser une ressource alternative se trouvent ainsi favorisés. Les individus capables d'exploiter une ressource secondaire, ont accès à une quantité moindre mais seront moins impactés par la compétition intraspécifique.
La compétition se fait alors en fonction du nombre d'individus capables d'utiliser la même ressource plutôt qu'en fonction de la taille totale de la population.
La compétition densité dépendante
Au sein d'une population, le nombre d'individus peut croître jusqu'au moment où le milieu atteint sa capacité de charge. Cette capacité de charge correspond à la densité maximale de population dans un milieu donné. Si la densité de population dépasse cette capacité de charge, elle agit alors de manière négative sur la fitness de chaque individu en réduisant la survie et/ou la reproduction. L'équilibre dans le milieu est atteint lorsque chaque individu peut produire un descendant viable.
Dans le cas de cette compétition, la relation entre la densité de la population et son taux de croissance est négative. En effet, plus la population va être importante, plus les interactions entre les individus seront conséquentes et agiront donc dans le but de freiner la croissance globale.
Réponses plastiques
Définition
Une réponse plastique est un changement au niveau du phénotype des individus, sans altérer les gènes qu'ils peuvent transmettre à leur descendance. Autrement dit c'est la capacité de chaque individu (ou d'une population) à s'adapter à un événement ponctuel sans que cette réponse se transmettre génétiquement à leur descendance.
Le phénotype est le résultat de l'expression des différents allèles d'un génotype. Cette expression est observable sous la forme des caractéristiques physiques et comportementales de l'individu pris en compte et varie au sein de la population.
D'après l'hypothèse de la variation de niche (Niche Variation Hypothesis en anglais), les variations de phénotype dans une population sont corrélées avec l'étendue de la niche écologique.
Stratégie optimale de recherche de nourriture
La théorie de la Stratégie optimale de recherche de la nourriture (Optimal Foraging Theory (OFT) en anglais) stipule que les prédateurs vont agir afin de maximiser leur apport d'énergie. Cette théorie permet d'expliquer pourquoi au sein d'une population à niche écologique alimentaire large, des individus peuvent choisir de se concentrer sur des proies différentes de la proie favorite de l'espèce quand la compétition intraspécifique augmente. En effet, la compétition va rendre la recherche de la proie favorite plus coûteuse en énergie et moins rentable que celle d'une autre proie plus facilement accessible.
Plusieurs scénarios prédisent que la compétition devrait conduire à de plus grandes variations entre individus. Cela se produit lorsque des individus, ayant des phénotypes différents, ont des proies favorites identiques mais peuvent exploiter des proies secondaires différentes lorsque leur proie principale devient rare. L'autre possibilité est que des variations du phénotype poussent des individus à s'écarter de la proie favorite plus souvent que les autres. Cela suggère que la compétition pour les ressources devrait conduire à des variations du régime alimentaire.
Certains de ces scénarios associent le fait que les variations du phénotype se font au niveau des changements du régime alimentaire en cas de forte compétition. Cette prédiction est analogue à la théorie évolutive de la diversification compétitive. La principale différence est que d'après le modèle de la stratégie optimale de recherche de nourriture, la compétition peut amener de la diversité par des changements dans le comportement de recherche de nourriture plutôt que des changements de la variabilité génétique.
Cependant les réponses plastiques et évolutives ne s'excluent pas mutuellement mais tendent plutôt à opérer sur des échelles de temps différentes. Il est possible que la diversification du comportement facilite la diversification évolutive.
Une étude réalisée sur une population d'épinoches par Richard Svanbäck et Daniel I. Bolnick a montré que le niveau de variation du régime alimentaire peut changer selon la compétition pour les ressources[5]. Il peut aussi se développer par le comportement, au lieu de la divergence évolutive étant donné que les résultats se sont vus au bout de deux semaines. C'est le temps nécessaire aux épinoches pour remarquer un changement dans la disponibilité des proies mais ce n'est pas suffisant pour qu'il y ait un changement génétique au fil des générations.
De telles diversifications comportementales peuvent apparaître beaucoup plus vite que des changements évolutifs et sont facilement réversibles.
Réponse au sein des colonies d'insectes eusociaux
Les insectes eusociaux ont un mode d'organisation en caste, avec une (ou plusieurs) femelles reproductives qui produisent des ouvrières non fertiles. En raison du très fort apparentement entre les individus de ces colonies (les ouvrières sont toutes sœurs au sein d'une colonie), il n'y a pas de conflits pour la reproduction et des stratégies de défense au niveau du groupe peuvent se mettre en place.
C'est ce qui a été démontré par Luc Passera, Eric Roncin, Bernard Kauffmann et Laurent Keller dans une étude sur la réponse des colonies de fourmis Pheidole pallidula à la compétition intraspécifique[6].
Il y a deux castes qui diffèrent dans leur morphologie au sein de ces colonies de fourmis : des ouvrières avec une petite tête qui s'occupent de la recherche de nourriture, des soins aux larves et pupes et du nettoyage de la fourmilière et des soldats avec une grande tête qui défendent la colonie contre les agresseurs.
Après avoir mis en contact visuel et phéromonal des colonies de Pheidole pallidula, il a été observé une plus grande production de soldats dans les colonies rencontrant de la compétition intraspécifique. À la fin de l'expérience, deux fois plus d'adultes soldats ont été observés dans les colonies ayant rencontré de la compétition intraspécifique que dans les colonies contrôles, mais sans changement de biomasse totale entre les deux colonies.
La différenciation, entre ouvrière et soldat, s'effectue au moment du développement de la larve en fonction de la nourriture qui lui est apporté. Ainsi la plus grande proportion de soldats observée, dans les colonies sous compétition, résulte d'un changement dans le comportement des ouvrières nourricières s'occupant des larves, après avoir perçu la présence de l'autre colonie.
Ces résultats démontrent un exemple notable de plasticité phénotypique en réponse à la compétition intraspécifique.
L'évolution de proportions flexibles entre soldats et ouvrières est possible car il n'y a pas de conflit entre la sélection individuelle et la sélection du groupe. En effet, ouvrières et soldats sont stériles et ne sont donc pas en compétition pour la reproduction.
Réponses évolutives
Définition
Une réponse évolutive est un changement au niveau de la sélection des génotypes, les plus avantageux vont ainsi se propager plus facilement dans la population.
On peut dire aussi qu'un certain phénotype, donnant un avantage non négligeable pour l'accès à la reproduction ou dans la survie, va être sélectionné. Il va donc être transmis plus facilement au sein de la population considérée.
Mécanismes
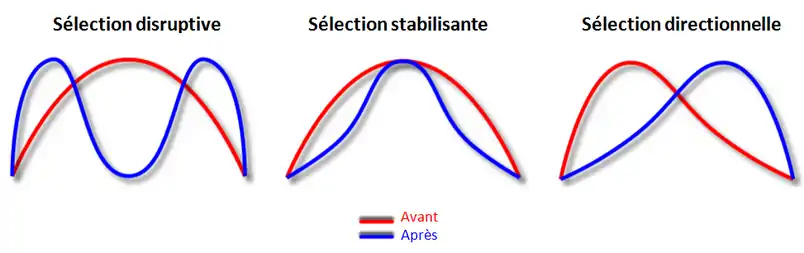
Il existe trois mécanismes de sélection :
- La sélection disruptive qui sélectionne les phénotypes les plus extrêmes dans la population et peut ainsi amener à une spéciation, c'est-à-dire la formation de deux espèces différentes par éloignement génétiques de deux groupes d'individus au sein de la même populations.
- La sélection stabilisante qui au contraire de la sélection disruptive favorise les phénotypes moyens et va donc réduire l'étendue de la niche écologique : les individus de la population vont devenir spécialistes c'est-à-dire qu'ils pourront utiliser peu de ressources différentes mais avec une très grande efficacité.
- La sélection directionnelle agit comme la sélection disruptive sauf qu'elle va favoriser uniquement un extrême de la gamme des phénotypes et va ainsi modifier le phénotype moyen de la population[7].
La spéciation compétitive
Quand une population est adaptée pour n'utiliser qu'une partie des ressources disponibles dans son environnement, la compétition fréquence-dépendante devrait favoriser l'expansion de la niche écologique.
C'est ce qui a été observé par Daniel I. Bolnick dans une étude sur la mouche Drosophila melanogaster[8] : il a placé des drosophiles non résistantes au cadmium dans des environnements contenant de la nourriture sans cadmium et de la nourriture avec cadmium. Il est parti du principe que les populations avec une forte compétition intraspécifique devraient évoluer plus vite pour tolérer le cadmium que les populations avec une faible compétition.
Pour cela, il a étudié l'évolution de la résistance des mouches au cadmium dans deux populations : l'une à faible compétition intraspécifique (LC) et l'autre à forte compétition (HC).
Les rares individus résistants au cadmium ont donc eu moins de compétition car ils étaient peu nombreux à pouvoir utiliser la nourriture au cadmium. Ils se sont, par conséquent, mieux reproduits et ont donné plus de descendance, ainsi les populations sont devenues résistantes au cadmium comme attendu par la théorie.
À partir de la quatrième génération, il a observé que le taux d'émergence des drosophiles résistantes au cadmium était significativement plus élevé dans la population à forte compétition que dans celle à faible compétition. Cette différence suggère qu'une compétition intense favorise les individus résistants au cadmium.
Il a ensuite réalisé des croisements, entre les mouches venant de la population à forte compétition et de celle à faible compétition. Il a observé que les descendants des individus de la population HC, s'étant développés sur de la nourriture riche en cadmium, ont une descendance plus résistante au cadmium que celle des individus de la même population, s'étant développés sur de la nourriture pauvre en cadmium. Ces derniers ont une descendance similaire à celle des individus de la population LC.
La population de drosophile à forte compétition a donc évolué rapidement pour utiliser le cadmium. Ce résultat confirme la théorie comme quoi la compétition intraspécifique peut favoriser la sélection d'une extension de la niche écologique et de la diversification des phénotypes liés.
Cette expérience correspond à la dynamique d'un modèle de spéciation compétitive, dans lequel une population utilisant seulement une partie de la ressource disponible a évolué pour utiliser l'ensemble de la ressource quand la compétition infraspécifique a augmenté.
La compétition favorise la diversification en baissant la valeur sélective des individus utilisant la ressource ancestrale, jusqu'au point où les ressources originellement moins optimales deviennent optimales pour une partie de la population ayant acquis la possibilité de les utiliser pour leur métabolisme.
Ce phénomène a déjà été vu dans d'autres espèces comme chez la mouche de la pomme (Rhagoletis pomonella), qui a une fécondité réduite sur les pommes comparé à sa ressource ancestrale : les fruits de l'aubépine. Cependant la compétition étant plus intense sur ces derniers, elle peut avoir poussé quelques populations à se spécialiser pour les pommes.
La sélection stabilisante
À l'inverse de la spéciation compétitive, la sélection stabilisante va favoriser le phénotype moyen et va uniformiser la population autour de ce phénotype[9].
C'est ce qu'ont démontré Christine Parent, Deepa Agashe et Daniel Bolnick dans trois expériences réalisées sur le scarabée Tribolium castaneum où ils ont mesuré comment la compétition modifie la taille de la niche écologique de la population au sein d'une génération[10] et après plusieurs générations :
Ils ont tout d'abord observé comment la valeur sélective des individus variait en fonction de la compétition intraspécifique sur trois types de ressources : une ressource constituée à 100 % de blé qui est la ressource ancestrale de cette espèce, une ressource constituée à 100 % de maïs qui est une nouvelle ressource pour l'espèce et une ressource donnant le choix aux scarabées entre du blé et du maïs.
Ils ont obtenu des résultats confirmant l'hypothèse que la compétition favoriserait l'expansion de la niche écologique : ils ont observé que la valeur sélective des individus était corrélée à la densité d'individus et donc à l'intensité de la compétition intraspécifique. Plus la compétition intraspécifique était élevée, plus la valeur sélective des individus était basse. Cependant, la valeur sélective des tribolium pouvant utiliser le blé ou le maïs était significativement plus élevée que celle des individus ne pouvant utiliser que le blé, qui eux-mêmes avaient une valeur sélective supérieure à celle des individus ne pouvant utiliser que le maïs.
Dans une deuxième expérience, ils ont déterminé quelles proportions de blé et de maïs confèrent la plus grande valeur sélective selon l'intensité de la compétition intraspécifique. La valeur sélective était calculée en regardant la longévité et la fécondité moyenne des individus.
Ils ont déterminé que la valeur sélective individuelle des tribolium est maximisée pour une proportion de 41 % de maïs lorsqu'il y a une faible compétition et pour 59 % de maïs quand il y a une forte compétition. Ces résultats montrent une proportion de maïs optimale autour de 50 % et confirment les résultats de la première expérience comme quoi un régime mixte est favorisé par la sélection et que la compétition favorise une plus grande consommation de maïs. Ces résultats montrent ainsi une estimation du paysage adaptatif de cette espèce.
Dans une troisième expérience, les chercheurs ont examiné l'effet de la compétition sur l'utilisation des ressources par les scarabées adultes ainsi que sur les ressources utilisées par leurs descendants.
Ils ont trouvé trois arguments montrant que la compétition réduit l'utilisation du maïs lorsque les scarabées peuvent choisir la composition de leur régime alimentaire :
Tout d'abord, ils ont trouvé que les scarabées adultes consomment environ 20 % de maïs lorsque la compétition est faible mais que cette proportion diminue lorsque la compétition augmente. Ce résultat contredit l'attente comme quoi la compétition devrait augmenter l'utilisation du maïs.
Ensuite, ils ont observé que les femelles préfèrent toujours pondre sur le maïs bien que le blé soit leur ressource ancestrale. Cependant, le taux de ponte sur le maïs diminue lorsque la compétition augmente. Enfin, ils ont vu que les larves venant de populations n'ayant jamais été en contact avec le maïs ont une préférence pour le blé accrue lorsque la compétition augmente. Une forte compétition intraspécifique provoque donc une contraction de la niche écologique des individus qui n'ont jamais connu la nouvelle ressource. En effet, ces individus n'ont eu qu'une ressource à exploiter et la sélection naturelle a donc favorisé ceux qui pouvaient l'exploiter le plus efficacement.
Ces trois résultats montrent tous que les tribolium utilisent plus le blé lorsque la compétition augmente, à la fois pour se nourrir et pour pondre. La diversité d'utilisation des ressources diminue donc avec l'augmentation de la compétition traduisant un effet de sélection stabilisante.
Comparaison avec d'autres forces évolutives
Les agents de sélection dans la nature, tels que la compétition interspécifique ou la prédation, jouent un rôle dans la formation du paysage adaptatif.
Comparaison avec la prédation
La compétition intraspécifique peut être un facteur de sélection plus fort que la prédation.
C'est ce qui a été montré par deux chercheurs, Ryan Calsbeek et Robert Cox, qui se sont intéressés aux populations de lézards Anolis sagrei des caraïbes et d’Amérique du Nord[11]. Les lézards vivant en Amérique sont peu nombreux et ont facilement accès à la nourriture qui est abondante, en revanche ils sont soumis à une pression importante de la part des prédateurs. Ceux vivant aux Caraïbes sont plus nombreux par unité de surface et sont donc en compétition pour la nourriture qui est la ressource limitante, cependant la prédation est beaucoup plus faible.
Ils ont reproduit expérimentalement sur des îlots entiers ce qu'ils observaient dans la nature. Ils ont appliqué trois traitements de prédations sur six îlots différents :
- deux îlots où la population de Anolis sagrei ne subit aucune prédation ;
- deux îlots où elle subit la prédation d'oiseaux uniquement ;
- deux îlots où elle subit une prédation dues aux oiseaux et aux serpents.
Ils ont aussi varié la densité d'anoles sur chaque îlot, chacun possédant entre 9 et 30 anoles pour 100 mètres carrés. Après avoir mesuré la taille du corps, des pattes et leur résistance à l'effort, ils ont marqué les anoles individuellement puis les ont relâchés sur les îlots.
Quelques mois plus tard, après la saison de reproduction, ils ont capturé les anoles survivants et ont reproduit les tests faits au départ. Ils ont montré que la prédation altère le comportement des lézards, qui montent plus haut dans les arbustes et qu'elle diminue leur survie. Cependant d'un point de vue morphologique, il n'y avait aucune modification. Alors que la longueur du corps et des pattes ainsi que l'endurance avaient augmenté chez les lézards soumis à la compétition intraspécifique. Ce changement morphologique s'explique par le fait que la taille des membres influence la rapidité et l'agilité des anoles, ce qui leur permet de mieux capturer leurs proies et de mieux échapper aux prédateurs. Ces traits permettent donc aux lézards de s'adapter face à la compétition, d'augmenter leur valeur sélective et ont ainsi été retenu par la sélection naturelle.
Cette expérience montre que la compétition intraspécifique est un puissant facteur de sélection qui sélectionne les morphologies les plus aptes à y faire face. Plus que la prédation, la compétition permet l'adaptation et l'évolution rapide des populations face aux contraintes de leur environnement.
Comparaison avec la compétition interspécifique
De même, la compétition intraspécifique peut être un agent de sélection plus fort que la compétition interspécifique.
Cette hypothèse a été observée chez deux espèces compétitives de poissons en Suède, la perche Perca fluviatilis et le gardon Rutilus rutilus[12]. Les chercheurs ont évalué les facteurs (compétition, prédation, et les facteurs abiotiques) qui pourraient influencer le choix de l'habitat (en zone pélagique ou en zone littorale) chez ces espèces compétitrices. La séparation pélagique/littorale dans des lacs inclut une séparation spatiale pour les ressources alimentaires puisque les différentes ressources sont associées aux habitats. En zone pélagique, la perche et le gardon rivalisent pour la ressource qui est le zooplancton et en zone littorale, les macro-invertébrés. Le gardon est un compétiteur supérieur par rapport à la perche pour le zooplancton pélagique alors que l'avantage compétitif est renversé dans la zone littorale où la perche est plus efficace que la gardon pour la recherche de nourriture.
Si la structure communautaire des poissons est essentiellement un résultat de la compétition interspécifique, la perche devrait alors se trouver principalement dans la zone littorale tandis que le gardon devrait occuper la zone pélagique. Si la compétition intraspécifique est plus forte que la compétition interspécifique, nous nous attendons à trouver le gardon et la perche dans les deux habitats, avec une différence de morphologie entre les individus du littoral et des zones pélagiques dans les deux espèces.
Les résultats ont montré que, peu importe la zone d'étude, on observe la présence aussi bien de perches et de gardons. Ils coexistent dans les deux habitats montrant une incompatibilité avec la compétition interspécifique, mais entrant dans le domaine de la compétition intraspécifique.
De plus, des variations morphologiques chez les poissons sont observées en fonction de l'habitat : la perche et le gardon attrapés dans la zone littorale ont un corps plus épais ainsi qu'une tête plus grande en comparaison de ceux attrapés dans la zone pélagique.
Le polymorphisme de ressource (variation morphologique associée à la variation dans l'utilisation de ressource) et le choix de l'habitat sont dus à des interactions intraspécifiques. En effet, la séparation morphologique est multi-espèces, ce qui montre que la compétition intraspécifique a eu un effet plus important que la compétition interspécifique sur la modification de la morphologie.
Notes et références
- (en) Charles Darwin, On the Origin of Species, London, England, Penguin Classics, , 516 p. (ISBN 9780140439120), p. 76
- (en) Michael Allaby, Scramble competition. A Dictionary of Zoology, 1999
- (en) J. MoyaLarano, M. Tigani El-Sayyid, C. Fox,Smaller beetles are better scramble competitors at cooler temperatures, Biol Lett. 22; 3(5), 2007 : 475-478.
- (en) Michael Allaby, Contest competition. A Dictionary of Zoology, 1999
- (en) Richard Svanbäck et Daniel I. Bolnick, « Intraspecific competition drives increased resource use diversity within a natural population », Proceedings of the Royal Society of London B: Biological Sciences, vol. 274, , p. 839-844 (ISSN 0962-8452 et 1471-2954, PMID 17251094, PMCID 2093969, DOI 10.1098/rspb.2006.0198, lire en ligne, consulté le )
- (en) Luc Passera, Eric Roncin, Bernard Kaufmann et Laurent Keller, « Increased soldier production in ant colonies exposed to intraspecific competition », Nature, no 379, , p. 360-361
- Kenneth Mather, « Polymorphism as an Outcome of Disruptive Selection », Evolution, vol. 9, , p. 52-61 (DOI 10.2307/2405357, lire en ligne, consulté le )
- (en) Daniel I. Bolnick, « Intraspecific competition favours niche width expansion in Drosophila melanogaster », Nature, no 410, , p. 463-466
- Kenneth Mather, « Polymorphism as an Outcome of Disruptive Selection », Evolution, vol. 9, , p. 52-61 (DOI 10.2307/2405357, lire en ligne, consulté le )
- (en) Christine E. Parent, Deepa Agashe et Daniel I. Bolnick, « Intraspecific competition reduces niche width in experimental populations », Ecology and Evolution, vol. 4, , p. 3978-3990 (ISSN 2045-7758, PMID 25505525, PMCID 4242580, DOI 10.1002/ece3.1254, lire en ligne, consulté le )
- (en) Ryan Calsbeek et Robert Cox, « Experimentally assessing the relative importance of predation and competition as agents of selection », Nature, no 465,
- (en) Richard Svanbäck, Peter Eklöv, Rebecka Fransson et Kerstin Holmgren, « Intraspecific competition drives multiple species resource polymorphism in fish communities », Oikos, no 117, , p. 114-124